

.gif)
¡BIENVENIDOS!
ESTE ES EL MARAVILLOSO MUNDO DE LA
BIOLOGÍA.
ESTE ES UN ESPACIO ESPECIALMENTE DISEÑADO PARA USTEDES. AQUÍ PODRÁN ENCONTRAR INFORMACIÓN MUY IMPORTANTE PARA ENRIQUECER SU CONOCIMIENTO. CON EL FIN DE APOYAR SUS PROCESOS ACADÉMICOS SE SUBIRÁ MATERIAL DIDÁCTICO Y, ADEMAS, SE DEJARAN ACTIVIDADES Y MUCHÍSIMOS RECURSOS MAS.
Maria Tereza Carmona duque
biology teacher
mariacarmona@windsorroyalschool.edu.co
SEGUNDO PERIODO
SEMANA DEL 17 AL 21 DE ABRIL
ESTE ES UN ESPACIO ESPECIALMENTE DISEÑADO PARA USTEDES. AQUÍ PODRÁN ENCONTRAR INFORMACIÓN MUY IMPORTANTE PARA ENRIQUECER SU CONOCIMIENTO. CON EL FIN DE APOYAR SUS PROCESOS ACADÉMICOS SE SUBIRÁ MATERIAL DIDÁCTICO Y, ADEMAS, SE DEJARAN ACTIVIDADES Y MUCHÍSIMOS RECURSOS MAS.
SEGUNDO PERIODO
SEMANA DEL 17 AL 21 DE ABRIL
TEMAS DE CUARTO PERIODO
CUARTO PERIODO
CUARTO PERIODO
Puntos más importantes:
-
Los productores, o autótrofos, fabrican su propias moléculas orgánicas. Los consumidores, o heterótrofos, obtienen las moléculas orgánicas al comer a otros organismos.
-
Una cadena alimenticia es una secuencia lineal de organismos a través de la cual los nutrientes y la energía pasan de un organismo a otro mediante consumo.
-
En una cadena alimenticia, cada organismo ocupa un nivel tróficodiferente, definido por cuántas transferencias de energía lo separan de la entrada en la base de la cadena.
-
Las redes tróficas consisten de varias cadenas alimenticias interconectadas y son una representación más realista de las relaciones de consumo en los ecosistemas.
-
La transferencia de energía entre niveles tróficos es ineficiente, con una eficiencia típica de alrededor del 10%. Esta ineficacia limita la longitud de las cadenas alimenticias.
- Los productores, o autótrofos, fabrican su propias moléculas orgánicas. Los consumidores, o heterótrofos, obtienen las moléculas orgánicas al comer a otros organismos.
- Una cadena alimenticia es una secuencia lineal de organismos a través de la cual los nutrientes y la energía pasan de un organismo a otro mediante consumo.
- En una cadena alimenticia, cada organismo ocupa un nivel tróficodiferente, definido por cuántas transferencias de energía lo separan de la entrada en la base de la cadena.
- Las redes tróficas consisten de varias cadenas alimenticias interconectadas y son una representación más realista de las relaciones de consumo en los ecosistemas.
- La transferencia de energía entre niveles tróficos es ineficiente, con una eficiencia típica de alrededor del 10%. Esta ineficacia limita la longitud de las cadenas alimenticias.
Introducción
Los organismos de diferentes especies interactúan de muchas maneras. Pueden competir o ser simbiontes, compañeros a largo plazo con una asociación muy cercana. Por supuesto, pueden hacer lo que a menudo vemos en los programas de la naturaleza: uno se come a otro: ¡ñam! Esto es, pueden formar uno de los eslabones de la cadena alimenticia.
En ecología, una cadena alimenticia es una serie de organismos que se comen entre ellos de forma que la energía y los nutrientes fluyan de uno al otro. Por ejemplo, si comiste una hamburguesa en el almuerzo, eres parte de una cadena alimenticia que se ve así: pasto vaca humano. Pero, ¿qué pasa si tu hamburguesa llevaba lechuga? En ese caso, también eres parte de una cadena alimenticia como esta: lechuga humano.
Como ilustra este ejemplo, no siempre podemos describir completamente lo que come un organismo, como el humano, mediante una vía lineal. Para situaciones como la de arriba, preferiríamos utilizar una red trófica, que está conformada por muchas cadenas alimenticias que se intersecan y que representan las diferentes cosas que un organismo puede comer, así como de qué otros organismos puede ser alimento.
En este artículo, estudiaremos más de cerca las cadenas alimenticias y redes tróficas para ver cómo representan el flujo de energía y nutrientes a través de los ecosistemas.
Los organismos de diferentes especies interactúan de muchas maneras. Pueden competir o ser simbiontes, compañeros a largo plazo con una asociación muy cercana. Por supuesto, pueden hacer lo que a menudo vemos en los programas de la naturaleza: uno se come a otro: ¡ñam! Esto es, pueden formar uno de los eslabones de la cadena alimenticia.
En ecología, una cadena alimenticia es una serie de organismos que se comen entre ellos de forma que la energía y los nutrientes fluyan de uno al otro. Por ejemplo, si comiste una hamburguesa en el almuerzo, eres parte de una cadena alimenticia que se ve así: pasto vaca humano. Pero, ¿qué pasa si tu hamburguesa llevaba lechuga? En ese caso, también eres parte de una cadena alimenticia como esta: lechuga humano.
Como ilustra este ejemplo, no siempre podemos describir completamente lo que come un organismo, como el humano, mediante una vía lineal. Para situaciones como la de arriba, preferiríamos utilizar una red trófica, que está conformada por muchas cadenas alimenticias que se intersecan y que representan las diferentes cosas que un organismo puede comer, así como de qué otros organismos puede ser alimento.
En este artículo, estudiaremos más de cerca las cadenas alimenticias y redes tróficas para ver cómo representan el flujo de energía y nutrientes a través de los ecosistemas.
Autótrofos contra heterótrofos
¿Qué estrategias básicas usan los organismos para obtener comida? Algunos organismos, llamados autótrofos, también conocidos como "los que se alimentan a sí mismos", pueden producir su propia comida, esto es, fabrican sus propios compuestos orgánicos a partir de moléculas sencillas como el dióxido de carbono. Hay dos tipos básicos de autótrofos:
-
Los fotoautótrofos, como las plantas, usan la energía solar para producir compuestos orgánicos —azúcares— a partir del dióxido de carbono mediante la fotosíntesis. Otros ejemplos de fotoautótrofos son las algas y las cianobacterias.
-
Los quimioautótrofos usan la energía de los químicos para producir compuestos orgánicos a partir de dióxido de carbono o moléculas similares. Este proceso se conoce como quimiosíntesis. Como ejemplo, hay bacterias quimioautótrofas que oxidan el ácido sulfhídrico que se encuentra en las comunidades de las fuentes hidrotermales en el fondo del mar, donde no llega la luz.
Los autótrofos son la base de todos los ecosistemas del planeta. Esto puede sonar muy dramático, ¡pero no es ninguna exageración! Los autótrofos forman la base de las cadenas alimenticias y las redes tróficas, y la energía que obtienen de la luz o los químicos sostiene a los demás organismos en la comunidad. Cuando hablamos de la función de los autótrofos dentro de las cadenas alimenticias, los llamamos productores.
Los heterótrofos, también conocidos como "los que se alimentan de otros", no pueden capturar la energía luminosa o química para fabricar su propia comida a partir de dióxido de carbono. Los humanos somos heterótrofos. Los heterótrofos obtenemos las moléculas orgánicas comiendo a otros organismos o sus productos. Los animales, los hongos y muchas bacterias son heterótrofos. Cuando hablamos de la función de los heterótrofos en las cadenas alimenticias, los llamamos consumidores. Como veremos enseguida, hay muchos tipos diferentes de consumidores con distintas funciones ecológicas, de los insectos que comen plantas, a los animales que comen carne, a los hongos que se alimentan de los residuos y desechos.
¿Qué estrategias básicas usan los organismos para obtener comida? Algunos organismos, llamados autótrofos, también conocidos como "los que se alimentan a sí mismos", pueden producir su propia comida, esto es, fabrican sus propios compuestos orgánicos a partir de moléculas sencillas como el dióxido de carbono. Hay dos tipos básicos de autótrofos:
- Los fotoautótrofos, como las plantas, usan la energía solar para producir compuestos orgánicos —azúcares— a partir del dióxido de carbono mediante la fotosíntesis. Otros ejemplos de fotoautótrofos son las algas y las cianobacterias.
- Los quimioautótrofos usan la energía de los químicos para producir compuestos orgánicos a partir de dióxido de carbono o moléculas similares. Este proceso se conoce como quimiosíntesis. Como ejemplo, hay bacterias quimioautótrofas que oxidan el ácido sulfhídrico que se encuentra en las comunidades de las fuentes hidrotermales en el fondo del mar, donde no llega la luz.
Los autótrofos son la base de todos los ecosistemas del planeta. Esto puede sonar muy dramático, ¡pero no es ninguna exageración! Los autótrofos forman la base de las cadenas alimenticias y las redes tróficas, y la energía que obtienen de la luz o los químicos sostiene a los demás organismos en la comunidad. Cuando hablamos de la función de los autótrofos dentro de las cadenas alimenticias, los llamamos productores.
Los heterótrofos, también conocidos como "los que se alimentan de otros", no pueden capturar la energía luminosa o química para fabricar su propia comida a partir de dióxido de carbono. Los humanos somos heterótrofos. Los heterótrofos obtenemos las moléculas orgánicas comiendo a otros organismos o sus productos. Los animales, los hongos y muchas bacterias son heterótrofos. Cuando hablamos de la función de los heterótrofos en las cadenas alimenticias, los llamamos consumidores. Como veremos enseguida, hay muchos tipos diferentes de consumidores con distintas funciones ecológicas, de los insectos que comen plantas, a los animales que comen carne, a los hongos que se alimentan de los residuos y desechos.
Cadenas alimenticias
Ahora, podemos echar un vistazo a cómo se mueven la energía y los nutrientes a través de una comunidad ecológica. Empecemos con algunas relaciones de quién se come a quién en una cadena alimenticia.
Una cadena alimenticia es una secuencia lineal de organismos a través de la cual la energía y los nutrientes se transfieren cuando un organismo se come a otro. Veamos las partes de una cadena alimenticia típica, comenzando desde la base —los productores— y moviéndonos hacia arriba.
-
En la base de la cadena alimenticia se encuentran los productores primarios. Los productores primarios son autótrofos y por lo general son plantas, algas o cianobacterias.
-
Los organismos que comen productores primarios se llaman consumidores primarios. Los consumidores primarios usualmente son herbívoros que comen plantas, aunque también pueden ser consumidores de algas o bacterias.
-
Los organismos que se comen a los consumidores primarios se llaman consumidores secundarios. Los consumidores secundarios por lo general comen carne: son carnívoros.
-
Los organismos que comen consumidores secundarios se llaman consumidores terciarios y son carnívoros que comen carnívoros, como las águilas o los peces grandes.
-
Algunas cadenas alimenticias tienen niveles adicionales, como los consumidores cuaternarios: carnívoros que comen consumidores terciarios. Los organismos que se encuentran hasta arriba en la cadena alimenticia se llaman superdepredadores.
Podemos ver ejemplos de estos niveles en el diagrama siguiente. Las algas verdes son productores primarios que son consumidas por moluscos, los consumidores primarios. Luego los moluscos se convierten en el almuerzo del Cottus cognatus, un pez que es un consumidor secundario y la comida de un pez más grande: el salmón real, un consumidor terciario.
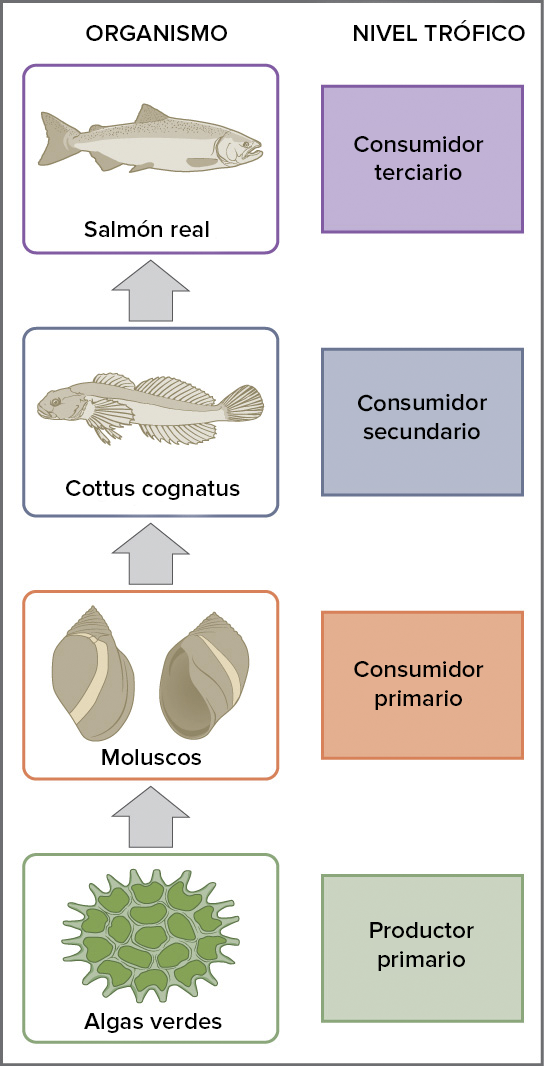 Crédito de imagen: Ecología de ecosistemas: Figura 3 por OpenStax College, Biology, CC BY 4.0
Cada una de las categorías anteriores se denomina nivel trófico y refleja cuántas transferencias de energía y nutrientes —cuántos pasos de consumo— separan a un organismo de la fuente original de la cadena alimenticia, la luz por ejemplo. Como veremos más adelante, asignar los organismos a los niveles tróficos no siempre es obvio; los humanos, por ejemplo, son omnívoros que pueden comer plantas y animales.
Crédito de imagen: Ecología de ecosistemas: Figura 3 por OpenStax College, Biology, CC BY 4.0
Cada una de las categorías anteriores se denomina nivel trófico y refleja cuántas transferencias de energía y nutrientes —cuántos pasos de consumo— separan a un organismo de la fuente original de la cadena alimenticia, la luz por ejemplo. Como veremos más adelante, asignar los organismos a los niveles tróficos no siempre es obvio; los humanos, por ejemplo, son omnívoros que pueden comer plantas y animales.
Ahora, podemos echar un vistazo a cómo se mueven la energía y los nutrientes a través de una comunidad ecológica. Empecemos con algunas relaciones de quién se come a quién en una cadena alimenticia.
Una cadena alimenticia es una secuencia lineal de organismos a través de la cual la energía y los nutrientes se transfieren cuando un organismo se come a otro. Veamos las partes de una cadena alimenticia típica, comenzando desde la base —los productores— y moviéndonos hacia arriba.
- En la base de la cadena alimenticia se encuentran los productores primarios. Los productores primarios son autótrofos y por lo general son plantas, algas o cianobacterias.
- Los organismos que comen productores primarios se llaman consumidores primarios. Los consumidores primarios usualmente son herbívoros que comen plantas, aunque también pueden ser consumidores de algas o bacterias.
- Los organismos que se comen a los consumidores primarios se llaman consumidores secundarios. Los consumidores secundarios por lo general comen carne: son carnívoros.
- Los organismos que comen consumidores secundarios se llaman consumidores terciarios y son carnívoros que comen carnívoros, como las águilas o los peces grandes.
- Algunas cadenas alimenticias tienen niveles adicionales, como los consumidores cuaternarios: carnívoros que comen consumidores terciarios. Los organismos que se encuentran hasta arriba en la cadena alimenticia se llaman superdepredadores.
Podemos ver ejemplos de estos niveles en el diagrama siguiente. Las algas verdes son productores primarios que son consumidas por moluscos, los consumidores primarios. Luego los moluscos se convierten en el almuerzo del Cottus cognatus, un pez que es un consumidor secundario y la comida de un pez más grande: el salmón real, un consumidor terciario.
Crédito de imagen: Ecología de ecosistemas: Figura 3 por OpenStax College, Biology, CC BY 4.0
Cada una de las categorías anteriores se denomina nivel trófico y refleja cuántas transferencias de energía y nutrientes —cuántos pasos de consumo— separan a un organismo de la fuente original de la cadena alimenticia, la luz por ejemplo. Como veremos más adelante, asignar los organismos a los niveles tróficos no siempre es obvio; los humanos, por ejemplo, son omnívoros que pueden comer plantas y animales.
Descomponedores
Hay otro grupo que vale la pena mencionar, aunque no siempre aparece en los diagramas de cadenas alimenticias. Este grupo es el de los descomponedores, organismos que degradan la materia orgánica muerta y los desechos.
A veces se considera a los descomponedores dentro de su propio nivel trófico. Como grupo, consumen la materia muerta y los productos de desecho que provienen de los demás niveles tróficos; por ejemplo, consumen materia vegetal en descomposición, el cuerpo a medio comer de una ardilla o los restos de un águila muerta. En cierto sentido, el nivel de los descomponedores es paralelo a los de la jerarquía estándar de los consumidores primarios, secundarios y terciarios.
Los hongos y bacterias son los descomponedores clave de muchos ecosistemas: usan la energía química en la materia muerta y los desechos para sus propios procesos metabólicos. Otros descomponedores son los detritívoros: consumidores de desechos y consumidores de residuos. Estos generalmente son animales multicelulares como las lombrices de tierra, los cangrejos, las babosas o los buitres. No solo se alimentan de materia orgánica muerta sino que la fragmentan también, poniéndola a disposición de las bacterias y los hongos descomponedores.
 Crédito de imagen: izquierda, Decomposers (Descomponedores) por Courtney Celley/USFWS, CC BY 2.0; derecha, Earthworm (Lombriz de tierra) por Luis Miguel Bugallo Sánchez, CC BY-SA 3.0
Los descomponedores como grupo juegan un papel crítico en el mantenimiento de la salud de los ecosistemas. Cuando descomponen la materia muerta y los desechos, liberan nutrientes que pueden ser reciclados y utilizados por los productores primarios.
Crédito de imagen: izquierda, Decomposers (Descomponedores) por Courtney Celley/USFWS, CC BY 2.0; derecha, Earthworm (Lombriz de tierra) por Luis Miguel Bugallo Sánchez, CC BY-SA 3.0
Los descomponedores como grupo juegan un papel crítico en el mantenimiento de la salud de los ecosistemas. Cuando descomponen la materia muerta y los desechos, liberan nutrientes que pueden ser reciclados y utilizados por los productores primarios.
Hay otro grupo que vale la pena mencionar, aunque no siempre aparece en los diagramas de cadenas alimenticias. Este grupo es el de los descomponedores, organismos que degradan la materia orgánica muerta y los desechos.
A veces se considera a los descomponedores dentro de su propio nivel trófico. Como grupo, consumen la materia muerta y los productos de desecho que provienen de los demás niveles tróficos; por ejemplo, consumen materia vegetal en descomposición, el cuerpo a medio comer de una ardilla o los restos de un águila muerta. En cierto sentido, el nivel de los descomponedores es paralelo a los de la jerarquía estándar de los consumidores primarios, secundarios y terciarios.
Los hongos y bacterias son los descomponedores clave de muchos ecosistemas: usan la energía química en la materia muerta y los desechos para sus propios procesos metabólicos. Otros descomponedores son los detritívoros: consumidores de desechos y consumidores de residuos. Estos generalmente son animales multicelulares como las lombrices de tierra, los cangrejos, las babosas o los buitres. No solo se alimentan de materia orgánica muerta sino que la fragmentan también, poniéndola a disposición de las bacterias y los hongos descomponedores.
Crédito de imagen: izquierda, Decomposers (Descomponedores) por Courtney Celley/USFWS, CC BY 2.0; derecha, Earthworm (Lombriz de tierra) por Luis Miguel Bugallo Sánchez, CC BY-SA 3.0
{kind=link}
Los descomponedores como grupo juegan un papel crítico en el mantenimiento de la salud de los ecosistemas. Cuando descomponen la materia muerta y los desechos, liberan nutrientes que pueden ser reciclados y utilizados por los productores primarios.
Redes tróficas
Las cadenas alimenticias nos dan una imagen clara de quién se come a quién. Sin embargo, surgen algunos problemas cuando tratamos de usarlas para describir comunidades ecológicas completas.
Por ejemplo, un organismo a veces puede comer muchos tipos de presa diferentes o ser consumido por varios depredadores, incluyendo aquellos que se encuentran en distintos niveles tróficos. ¡Eso es lo que sucede cuando te comes una hamburguesa! La vaca es un consumidor primario y la hoja de lechuga es un productor primario.
Para representar estas relaciones con más precisión, podemos usar una red trófica, una gráfica que muestre todas las interacciones tróficas (asociadas a la alimentación) entre las diferentes especies de un ecosistema. El diagrama de abajo muestra un ejemplo de una red trófica del Lago Ontario. Los productores primarios están marcados en verde, los consumidores primarios en naranja, los consumidores secundarios en azul y los consumidores terciarios en morado.
 Crédito de imagen: Ecología de ecosistemas: Figura 5 por OpenStax College, Biology, CC BY 4.0; obra original del NOAA, GLERL
En las redes tróficas, las flechas apuntan desde un organismo que es devorado hacia el que se lo come. Como muestra la red trófica de arriba, algunas especies pueden comer organismos de más de un nivel trófico. Por ejemplo, los camarones misidáceos comen tanto productores primarios como consumidores primarios.
Pregunta extra: esta red trófica tiene una cadena alimenticia que vimos anteriormente en el artículo: algas verdes moluscos el pecesito Cottus cognatus salmón real. ¿Puedes encontrarla?
Crédito de imagen: Ecología de ecosistemas: Figura 5 por OpenStax College, Biology, CC BY 4.0; obra original del NOAA, GLERL
En las redes tróficas, las flechas apuntan desde un organismo que es devorado hacia el que se lo come. Como muestra la red trófica de arriba, algunas especies pueden comer organismos de más de un nivel trófico. Por ejemplo, los camarones misidáceos comen tanto productores primarios como consumidores primarios.
Pregunta extra: esta red trófica tiene una cadena alimenticia que vimos anteriormente en el artículo: algas verdes moluscos el pecesito Cottus cognatus salmón real. ¿Puedes encontrarla?
Las cadenas alimenticias nos dan una imagen clara de quién se come a quién. Sin embargo, surgen algunos problemas cuando tratamos de usarlas para describir comunidades ecológicas completas.
Por ejemplo, un organismo a veces puede comer muchos tipos de presa diferentes o ser consumido por varios depredadores, incluyendo aquellos que se encuentran en distintos niveles tróficos. ¡Eso es lo que sucede cuando te comes una hamburguesa! La vaca es un consumidor primario y la hoja de lechuga es un productor primario.
Para representar estas relaciones con más precisión, podemos usar una red trófica, una gráfica que muestre todas las interacciones tróficas (asociadas a la alimentación) entre las diferentes especies de un ecosistema. El diagrama de abajo muestra un ejemplo de una red trófica del Lago Ontario. Los productores primarios están marcados en verde, los consumidores primarios en naranja, los consumidores secundarios en azul y los consumidores terciarios en morado.
Crédito de imagen: Ecología de ecosistemas: Figura 5 por OpenStax College, Biology, CC BY 4.0; obra original del NOAA, GLERL
En las redes tróficas, las flechas apuntan desde un organismo que es devorado hacia el que se lo come. Como muestra la red trófica de arriba, algunas especies pueden comer organismos de más de un nivel trófico. Por ejemplo, los camarones misidáceos comen tanto productores primarios como consumidores primarios.
Pregunta extra: esta red trófica tiene una cadena alimenticia que vimos anteriormente en el artículo: algas verdes moluscos el pecesito Cottus cognatus salmón real. ¿Puedes encontrarla?
Redes tróficas de pastoreo contra redes tróficas de detritos
Las redes tróficas normalmente no muestran a los descomponedores, puede que hayas notado que la red trófica del Lago Ontario que vimos anteriormente no lo hace. Aún así, todos los ecosistemas necesitan formas de reciclar la materia muerta y los desechos. Esto significa que los descomponedores en realidad están ahí, aunque no se los mencione mucho.
Por ejemplo, en el ecosistema de pradera que se muestra abajo, hay una red trófica de pastoreo de plantas y animales que alimentan la red trófica de detritos de bacterias, hongos y detritívoros. La red de detritos se muestra en forma simplificada en la banda café que se encuentra en la parte inferior del diagrama. En realidad, consiste de varias especies relacionadas mediante interacciones de alimentación específicas, conectadas por flechas, como sucede con la red de pastoreo en la parte superior. Las redes de detritos pueden proveer de energía a las redes de pastoreo, como cuando un petirrojo come una lombriz de tierra.
 Crédito de imagen: modificada de El flujo de energía a través de los ecosistemas: Figura 5 por OpenStax College, Biology, CC BY 4.0; para ver los créditos de imagen completos de las imágenes originales, consulta la ventana emergente que sigue.
Crédito de imagen: modificada de El flujo de energía a través de los ecosistemas: Figura 5 por OpenStax College, Biology, CC BY 4.0; para ver los créditos de imagen completos de las imágenes originales, consulta la ventana emergente que sigue.
Las redes tróficas normalmente no muestran a los descomponedores, puede que hayas notado que la red trófica del Lago Ontario que vimos anteriormente no lo hace. Aún así, todos los ecosistemas necesitan formas de reciclar la materia muerta y los desechos. Esto significa que los descomponedores en realidad están ahí, aunque no se los mencione mucho.
Por ejemplo, en el ecosistema de pradera que se muestra abajo, hay una red trófica de pastoreo de plantas y animales que alimentan la red trófica de detritos de bacterias, hongos y detritívoros. La red de detritos se muestra en forma simplificada en la banda café que se encuentra en la parte inferior del diagrama. En realidad, consiste de varias especies relacionadas mediante interacciones de alimentación específicas, conectadas por flechas, como sucede con la red de pastoreo en la parte superior. Las redes de detritos pueden proveer de energía a las redes de pastoreo, como cuando un petirrojo come una lombriz de tierra.
Crédito de imagen: modificada de El flujo de energía a través de los ecosistemas: Figura 5 por OpenStax College, Biology, CC BY 4.0; para ver los créditos de imagen completos de las imágenes originales, consulta la ventana emergente que sigue.
La eficiencia en la transferencia de energía limita la longitud de las cadenas alimenticias
La energía se transfiere entre los niveles tróficos cuando un organismo se come a otro y obtiene las moléculas ricas en energía del cuerpo de su presa. Sin embargo, esta transferencia es ineficiente y esta ineficacia limita la longitud de las cadenas alimentarias.
Cuando la energía entra en un nivel trófico, parte de ella es almacenada como biomasa, pasa a formar parte del cuerpo del organismo. Esta es la energía que queda disponible para el siguiente nivel trófico, ya que solo la energía almacenada como biomasa puede ser consumida. Por regla general, solo alrededor del 10% de la energía almacenada como biomasa en un nivel trófico, por unidad de tiempo, termina como biomasa en el siguiente nivel trófico, en la misma unidad de tiempo. Es bueno tener en mente esta regla del 10% de transferencia de energía.
Como ejemplo, supongamos que los productores primarios de un ecosistema almacenan 20,000 kcal/m/año de energía en biomasa. Esta es también la cantidad de energía disponible por año para los consumidores primarios que se comen a los productores. La regla del 10% predice que los consumidores primarios solo almacenarán 2,000 kcal/m/año de energía en sus cuerpos, lo que pone la energía a disposición de los depredadores —consumidores secundarios— a una tasa menor.
Este patrón de transferencia parcial limita la longitud de las cadenas alimenticias; después de cierto número de niveles tróficos, por lo general entre tres y seis, queda muy poco flujo de energía para mantener una población en un nivel superior.
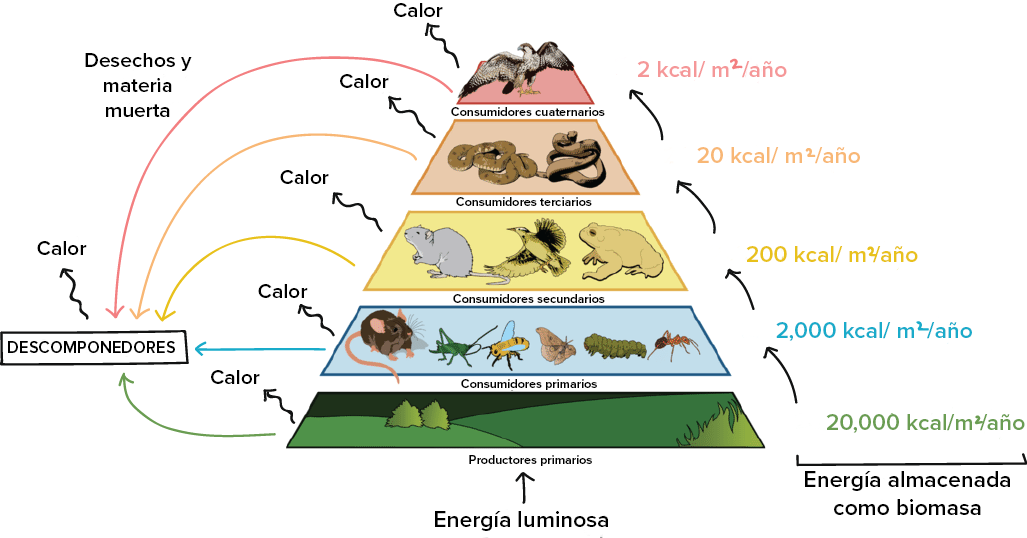 Crédito de imagen: modificación de Ecological pyramid (Pirámide ecológica) por CK-12 Foundation, CC BY-NC 3.0
¿Por qué sale tanta energía entre un nivel trófico y el siguiente? Estas son algunas de las razones principales que explican la transferencia de energía ineficiente:
Crédito de imagen: modificación de Ecological pyramid (Pirámide ecológica) por CK-12 Foundation, CC BY-NC 3.0
¿Por qué sale tanta energía entre un nivel trófico y el siguiente? Estas son algunas de las razones principales que explican la transferencia de energía ineficiente:
-
En cada nivel trófico, una cantidad significativa de energía se disipa como calor a medida que los organismos llevan a cabo la respiración celular y realizan sus vidas diarias.
-
Parte de las moléculas orgánicas que consume un organismo no son digeridas y salen del cuerpo como heces, excrementos, en lugar de ser utilizadas.
-
No todos los organismos individuales en un nivel trófico serán devorados por los organismos del siguiente nivel, algunos morirán sin haber sido consumidos.
Las heces y los organismos muertos no consumidos se convierten en alimento para los descomponedores, quienes los metabolizan y convierten su energía en calor mediante la respiración celular. Así que, la energía no desaparece en realidad, al final toda termina como calor.
La energía se transfiere entre los niveles tróficos cuando un organismo se come a otro y obtiene las moléculas ricas en energía del cuerpo de su presa. Sin embargo, esta transferencia es ineficiente y esta ineficacia limita la longitud de las cadenas alimentarias.
Cuando la energía entra en un nivel trófico, parte de ella es almacenada como biomasa, pasa a formar parte del cuerpo del organismo. Esta es la energía que queda disponible para el siguiente nivel trófico, ya que solo la energía almacenada como biomasa puede ser consumida. Por regla general, solo alrededor del 10% de la energía almacenada como biomasa en un nivel trófico, por unidad de tiempo, termina como biomasa en el siguiente nivel trófico, en la misma unidad de tiempo. Es bueno tener en mente esta regla del 10% de transferencia de energía.
Como ejemplo, supongamos que los productores primarios de un ecosistema almacenan 20,000 kcal/m/año de energía en biomasa. Esta es también la cantidad de energía disponible por año para los consumidores primarios que se comen a los productores. La regla del 10% predice que los consumidores primarios solo almacenarán 2,000 kcal/m/año de energía en sus cuerpos, lo que pone la energía a disposición de los depredadores —consumidores secundarios— a una tasa menor.
Este patrón de transferencia parcial limita la longitud de las cadenas alimenticias; después de cierto número de niveles tróficos, por lo general entre tres y seis, queda muy poco flujo de energía para mantener una población en un nivel superior.
Crédito de imagen: modificación de Ecological pyramid (Pirámide ecológica) por CK-12 Foundation, CC BY-NC 3.0
¿Por qué sale tanta energía entre un nivel trófico y el siguiente? Estas son algunas de las razones principales que explican la transferencia de energía ineficiente:
- En cada nivel trófico, una cantidad significativa de energía se disipa como calor a medida que los organismos llevan a cabo la respiración celular y realizan sus vidas diarias.
- Parte de las moléculas orgánicas que consume un organismo no son digeridas y salen del cuerpo como heces, excrementos, en lugar de ser utilizadas.
- No todos los organismos individuales en un nivel trófico serán devorados por los organismos del siguiente nivel, algunos morirán sin haber sido consumidos.
Las heces y los organismos muertos no consumidos se convierten en alimento para los descomponedores, quienes los metabolizan y convierten su energía en calor mediante la respiración celular. Así que, la energía no desaparece en realidad, al final toda termina como calor.
TEMAS DE TERCER PERIODO
TALLER FINAL PARA ENTREGAR EL DÍA 17 DE AGOSTO
EN EL CUADERNO
REPRODUCCIÓN HUMANA
Seré exigente con los dibujos y diagramas
TEMAS DEL 1 AL 12 DE AGOSTO
Todos los seres vivos tienen la capacidad de producir nuevos individuos. Esa es una característica fundamental de los organismos para la continuidad de la especie y de la vida. Esta capacidad no es otra cosa que la biogénesis, o lo que comúnmente llamamos reproducción. Existen dos formas de reproducción: la sexual y la asexual. En este artículo se expondrá únicamente la reproducción asexual en los animales.

Este tipo de reproducción puede llevarse a cabo por:
Ventajas de la reproducción asexual
SEMANA DEL 25 AL 29 DE JULIO
ESCRIBE LAS SIGUIENTES PREGUNTAS!
RESULTADOS
SEMANA DEL 11 AL 15 DE JULIO
SEMANA DEL 5 AL 8 DE JULIO
La reproducción asexual o asexualidad involucra un tipo de reproducción donde la constitución genética de los dos organismos que se producen es idéntica a la del progenitor.
EN EL CUADERNO
REPRODUCCIÓN HUMANA
Seré exigente con los dibujos y diagramas
- ¿Que es la menstruación y que se expulsa en ella?
- ¿En que parte del aparato reproductor femenino ocurre la fecundación?
- ¿Cuales son las gónadas femeninas y cuales las masculinas?
- ¿Cuales son los gametos femeninos y masculinos?
- ¿Cuales son las partes del espermatozoide Dibújalas y explica la función de cada una
- ¿Que nombre recibe la capa del útero donde se implantara el embrión ?
- ¿Que función sexual reproductiva cumplen las feromonas?
- ¿Cual es la hormona que regula los caracteres sexuales masculinos?
- ¿Cual es la hormona que regula los caracteres sexuales femeninos?
- ¿Que función cumplen los estrogenos y como actúan en la anticoncepción?
- ¿Como y cuando ocurre la ovulación y?
- Dibuja los aparatos reproductores femeninos y masculinos con sus respectivas partes.
TEMAS DEL 1 AL 12 DE AGOSTO
La reproducción sexual en animales
Todos los organismos animales proceden del desarrollo de una célula llamada huevo o cigoto que procede de la unión de los gametos, un espermatozoide y un óvulo, células especiales que se forman en las gónadas, testículos y ovarios respectivamente.
En los animales existe una gran diversidad de formas de reproducción sexual, la mayoría son unisexuales pero algunos grupos son hermafroditas como determinados anélidos, moluscos o peces, realizando una fecundación cruzada entre dos organismos, ya que no se pueden fecundar a sí mismos.
ENLACE IMPORTANTE
La reproducción asexual en los animales
 |
| Animales sésiles que se reproducen asexualmente |
Tipos de reproducción asexual
La reproducción asexual ocurre sin la intervención de órganos sexuales especiales, produciéndose nuevos individuos a partir de un solo progenitor. En las plantas se presenta mediante la producción de esporas o bulbos. En los animales protistas, en gusanos poliquetos y planarias, y en animales sésiles acuáticos como los tunicados, cnidarios y esponjas la reproducción ocurre de distintas maneras.

Este tipo de reproducción puede llevarse a cabo por:
- Escisión unicelular - un animal se divide en mitades generalmente iguales que se desarrollan hasta formar dos adultos idénticos al original. En este proceso de división primero se divide el núcleo y luego el citoplasma. Los esporozoos, como el Paramecio, llevan a cabo este tipo de reproducción.
 |
| Paramecio en proceso de división |
- Esporulación - en este caso el núcleo se divide varias veces y el citoplasma también. Cada pedazo de núcleo se lleva una porción del citoplasma. De cada división surge un nuevo individuo.
- Gemación - en este tipo de reproducción los nuevos individuos nacen en forma de yema, adheridos al original, formando una colonia. Ejemplos de animales que se reproducen así son: los tunicados y cnidarios como la hidra.
 |
| Por gemación la hidra ha creado un nuevo individuo. |
- Fragmentación - esta forma de reproducción es algo curiosa. El individuo original se divide en varias partes y de cada parte surge un individuo nuevo idéntico al original. Algunos animales en los que se da este proceso son los nemertinos (gusanos) y turbelarios (planarias o gusanos planos).
 |
| Nacimiento de una nueva estrella de mar por el proceso de fragmentación |
 |
| De este gusano partido en dos partes surgirán dos gusanos iguales al original, regenerando la parte faltante. |
Ventajas de la reproducción asexual
- No necesita de una pareja para llevarse a cabo la reproducción.
- No requieren de una madurez sexual que los animales más evolucionados si necesitan.
- La reproducción puede ocurrir en cualquier momento.
- Es un proceso simple y sencillo.
- Se puede multiplicar la población rápidamente.
- Un individuo tiene mayor oportunidad de supervivencia de la especie de quedar aislado.
Desventajas de la reproducción asexual
- No hay un aporte genético de dos padres, por lo tanto el resultado es... copias iguales al original. Esto implica que no hay un aporte de información genética nueva para la evolución de los nuevos individuos.
Sea sexual o asexual la reproducción de los animales, es un proceso de importancia vital para su supervivencia
SEMANA DEL 25 AL 29 DE JULIO
ESCRIBE LAS SIGUIENTES PREGUNTAS!
1) ¿ Cómo es la reproducción asexual o vegetativa de las plantas?
2) ¿ Cuándo se da la reproducción sexual en las plantas?
3) ¿ Cómo ocurre la polinización.?
4) ¿ Cúales son los animales encargados de la polinización?
5) ¿ Qué son los estambres y los pistilos en las flores?
6) ¿ Por que se le da el nombre de hermafroditas a algunas flores?
7) Realiza los dibujos correspondientes
8) ¿Que significa que una planta es dioica o monoica?
RESULTADOS
ESTUDIANTES QUE NO DEBEN PRESENTAR ACTIVIDADES DE NIVELACIÓN PARA EL SEGUNDO PERIODO
MIGUEL FUENTES
JOSE PADILLA
JUAN DIEGO
JUAN CAMILO SALAZAR
DANIELA VILLALBA
LOS DEMÁS DEBEN PRESENTAR LA SIGUIENTE ACTIVIDAD DE NIVELACIÓN: EVALUACIÓN DE LOS TEMAS VISTOS EN EL SEGUNDO PERIODO
DÍA DEL EXAMEN JULIO 12 DEL 2016
Reproducción asexual de las plantas
Existen dos tipos de reproducción asexual en plantas: multiplicación vegetativa y por gérmenes:
1. Multiplicación vegetativa: Asegura la perpetuación de individuos bien adaptados a ese medio y evolutivamente eficaces. Es muy común incluso en plantas superiores. Existen dos tipos: la fragmentación y la división celular que engloba la bipartición y la gemación.
La fragmentación consiste la fragmentación de partes de células, talos o vástagos de los que surgen individuos hijos. Un ejemplo de fragmentación son los esquejes. En la bipartición, la célula madre se divide por completo en dos células hijas nuevas de igual tamaño. En la gemacióncelular el tamaño de la célula hija es al principio menor que el de la célula madre.
2. Por gérmenes. Los gérmenes son células asexuales reproductivas que desarrollan directamente el individuo. Existen varios tipos: pluricelulares -los propágulos- y generalmente unicelulares -las esporas-.
Hay zonas en que porciones del talo o del tallo de las plantas pluricelulares están particularmente especializadas para separarse de la planta madre y extenderse, son los propágulos (agrupaciones de células), son muy comunes en las plantas inferiores. Existen varios tipos, los hormogonios de las cianobacterias, los tubérculos de la patata, los dientes del ajo…
Las esporas son células germinales especialmente diferenciadas para la reproducción asexual.
Esporas
Son la forma más corriente de reproducción asexual en plantas, producen en general poca variabilidad, son agentes de dispersión y normalmente unicelulareas aunque hay esporas con varias células o núcleos.
Existen varios tipos de esporas según las condiciones de formación:
- Según la situación: exósporas o conidios si se forman al exterior por estrangulación y endósporas si se forman en el interior de un esporangio.
- Según la capacidad de dispersión: aplanósporas si son inmóviles como el polen, muchosconidios y zoosporas o planósporas si son móviles.
- Según la formación: mitósporas o neutrósporas si son diploide y meiosporas, gonosporas o esporas “sexuales” si son haploides.
Las esporas tienen también nombres especiales como por ejemplo: diplosporas si son diploide, haplósporas si son haploides, si son esporas de resistencia se las llama clamidosporas. Si se producen en ascas son ascosporas, basidiosporas si se producen en basidios. Heterósporas si son distintas generalmente de tamaño, micrósporas si son pequeñas y masculinas, megásporas si son grandes y femeninas.
Las estructuras especializadas donde se producen las esporas son los esporangios. Son unicelulares (sin cubierta) en algas y hongos; pluricelulares (con cubierta y arquesporio que es el tejido fértil) de briófitos a espermatófitos. Su nomenclatura es igual que la de las esporas, por ejemplo de meióspora meiosporangio.
Reproducción sexual
Este tipo de reproducción es un mecanismo mucho más complicado que la reproducción asexual y en ella intervienen muchos factores.
La reproducción sexual implica la unión de células germinales especiales, los gametos, y está encaminada a la variabilidad genética por recombinación cromosómica. Este proceso se realiza en varias etapas. Primero se realiza la meiosis para transformar las células diploides en haploides que son los gametos.
Posteriormente se produce la singamia o unión de gametos haploides para formar un zigoto diploide, que implica una plasmogamia (unión de citoplasmas) y una cariogamia o fecundación (unión de núcleos).
Los gametos suelen ser haploides, (n), y de polaridades (sexos) opuestos, además se producen en estructuras especiales, los gametangios.
Existen varios tipos de reproducción sexual:
- Isogamia: unión de gametos de igual forma y tamaño pero de polaridades distintas.
- Anisogamia: Unión de gametos distintos en forma y tamaño y de polaridad opuesta.
- Oogamia: es un caso especial de anisogamia pero con un gameto femenino inmóvil y de mayor tamaño que el masculino.
Los gametos, al igual que las esporas, reciben distintos nombres. Los gametos femeninos se llaman ovocélula, oosfera, óvulo; y los masculinos anterozoide, anterozoo, espermatozoide, espermatozoo, espermacio que puede ser inmóvil en algunos hongos.
Los gametangios también reciben nombres especiales, existen los mismos tipos que los de los esporangios. Los gametangios primitivos son unicelulares y pueden estar recubiertos por una o varias paredes celulares.
Los gametangios más evolucionados son los pluricelulares con cubierta de protección. En la oogamia, el gametangio masculino se llama anteridio y posee un menor tamaño y forma filiforme, el femenino se llama oogonio y tiene mayor tamaño y forma esférica. De Briófitos en adelante, al masculino se le llama anteridio y al femenino arquegonio.
Existen casos especiales de gametangiogamia en la que la fecundación se lleva a cabo por fusión de gametangios y posterior unión de gametos; de somatogamia en la que no se producen gametangios y se copulan células somáticas; y apomixis, reproducción sexual sin fecundación, las células somáticas hacen el papel de germinales. En este artículo puedes encontrar más información sobre cómo se forman los espermatozoides de las plantas.
La reproducción de las plantas angiospermas
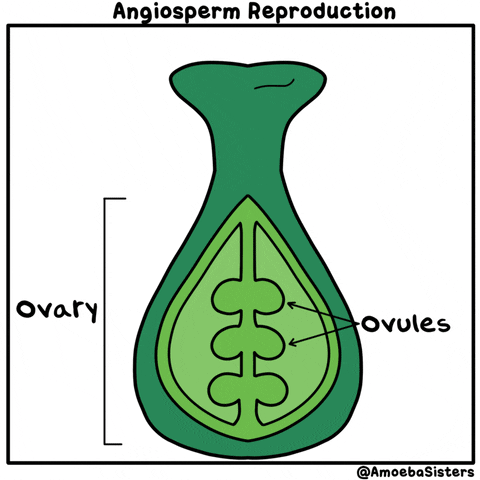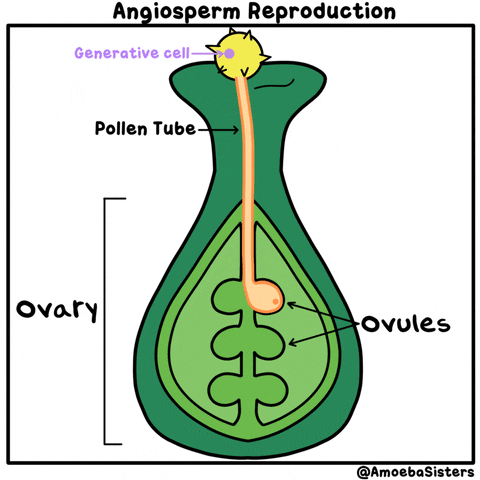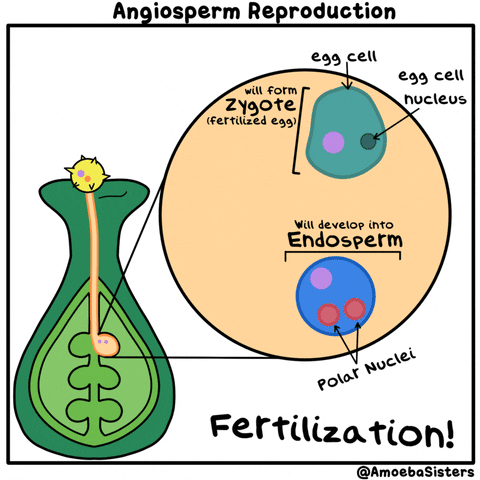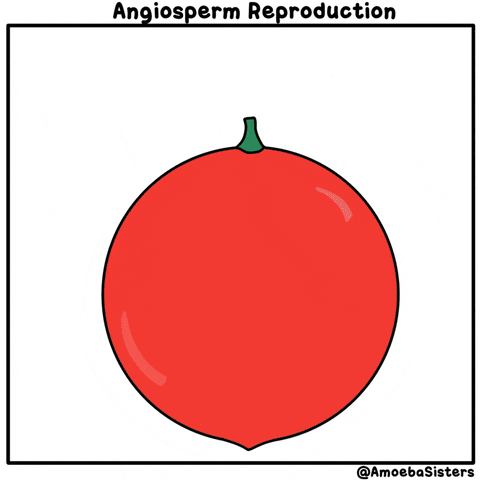
La reproducción sexual de las plantas angiospermas es el ejemplo más popular. Es la reproducción de las plantas con flores. Cuando un grano de polen llega al pistilo de la flor, se produce la polinización. Al entrar en su interior, se produce la generación de las células germinales o gametos masculinos que fecundan los óvulos que hay en el gineceo mediante el tubo polinizador.
Una vez los óvulos están fecundados, proceso conocido como fertilización, se forman las semillas, y una vez formadas éstas, el gineceo se convierte en el fruto que habitualmente conocemos como resultado de la reproducción de las plantas.
En la fecundación se produce la unión dos células haploides con distinto material genético, que se unen formando un individuo de genotipo diferente del de los padres y por este motivo se llama reproducción sexual.
Origen y evolución de la sexualidad
Se piensa que los eucariotas surgieron hace unos 600 M .A. a partir de procariotas debido a la invasión de nuevos hábitats y que estos eucariotas primitivos son los protoctistas.
La aparición de la reproducción sexual se produjo a partir del perfeccionamiento de la mitosis y la aparición de la meiosis y ciclo sexual. El origen de la mitosis pudo producirse en los protoctistas y especialmente en los dinoflagelados ya que su mitosis es “anómala”, no hay huso mitótico.
El origen de la reproducción sexual también pudo haberse dado en protoctistas al mismo tiempo que la mitosis debido a que estos organismos poseen ambos procesos incompletos. Surgió como consecuencia de la ventaja adaptativa que suponía ya que al haber recombinación cromosómica es exploradora e innovadora y facilita la colonización de nuevos hábitats. En la reproduccion de las plantas se da este tipo como ya hemos visto junto con la reproduccion asexual.
Si quieres ampliar más información sobre la reproducción sexual y asexual de las plantas o la botánica en general, te recomendamos el libro Strasburger: Tratado de botánica del que se ha extraído parte de esta información. Además, es el libro clásico y de cabecera de cualquier estudiante de botánica.
SEMANA DEL 5 AL 8 DE JULIO
TIPOS DE REPRODUCCIÓN ASEXUAL
La reproducción asexual o asexualidad involucra un tipo de reproducción donde la constitución genética de los dos organismos que se producen es idéntica a la del progenitor.
Existen distintos tipos de reproducción asexual entre los que se encuentran la bipartición o simple fisión, la gemación, la fragmentación y la esporulación.
• La bipartición o fisión, es un tipo de reproducción asexual en el que la célula se divide dando origen a dos células genéticamente idénticas entre sí pero de menor tamaño que la inicial. Esta modalidad de reproducción se da principalmente en organismos unicelulares como los protozoos y las bacterias.
En las bacterias, el proceso se denomina fisión binaria transversal, y se caracteriza porque el material hereditario (DNA) de la bacteria, se duplica y luego se separa; y la pared celular se forma en el interior y en sentido transversal.
• La gemación ocurre en organismos unicelulares y multicelulares. En la superficie del progenitor se forma una gema que crece y se estrangula hasta separarse por completo del organismo original. Las células producidas pueden tener vida propia o formar colonias si permanecen unidas al organismo parental. La levadura es un hongo que se reproduce por gemación. La gemación también ocurre en organismos pluricelulares. Un ejemplo de esto lo constituye la hidra. Sobre la superficie corporal de este organismo pluricelular, aparecen abultamien-tos que luego darán origen a nuevas hidras, genéticamente idénticas a su progenitor, pero de menor tamaño.
• La fragmentación. En este tipo de reproducción asexual se origina un nuevo organismo a partir de fragmentos del organismo progenitor. Ésta se da en organismos como las plenarias, que cuando se cortan por la mitad originan un nuevo organismo a partir de cada mitad.
A medida que se asciende en la escala zoológica, esta modalidad de reproducción va desapareciendo, y es reemplazada por mecanismos de regeneración de tejidos dañados. Por ejemplo, una lagartija (Leo-laemus montícola) puede regenerar parte de su cola cuando la ha perdido, la estrella de mar regenera parte del brazo perdido; sin embargo, el hombre tiene menos facilidad para regenerar estructuras dañadas: el crecimiento de uñas, del cabello y la cicatrización son algunos ejemplos
Tercera Ley De Mendel
La tercera ley de Mendel, también llamada Ley de la Herencia Independiente de Caracteres o Ley de la Asociación Independiente. Según Mendel, hay rasgos heredados que se obtienen de forma independiente, sin relación con el fenotipo, lo cual no afecta al patrón de herencia de otros rasgos. Esta ley se cumple en los genes que no están ligados, es decir que se encuentran en diferentes cromosomas o que están en zonas muy separadas del mismo cromosoma.
Mendel, para concluir la tercera de las leyes de Mendel, realizó un cruce de plantas de chícharos que producían semillas amarillas y llanas, con chícharos que producían semillas verdes y con textura irregular. Estas eran homocigóticas para los dos caracteres de textura y color. Se concluía que la ley de uniformidad estaba presente, pues con la primera generación se pudo obtener semillas amarillas y lisas.
 Sin embargo, al cruzar esta primera generación para obtener una segunda generación, se observan nuevos tipos de semillas con caracteres diversos pero relacionados con la generación parental, se obtuvieron semillas amarillas y lisas, amarillas y rugosas, verdes y lisas, y verdes y rugosas. Sigue leyendo aquí para aprender más acerca de Tercera Ley De Mendel.
Sin embargo, al cruzar esta primera generación para obtener una segunda generación, se observan nuevos tipos de semillas con caracteres diversos pero relacionados con la generación parental, se obtuvieron semillas amarillas y lisas, amarillas y rugosas, verdes y lisas, y verdes y rugosas. Sigue leyendo aquí para aprender más acerca de Tercera Ley De Mendel.
La tercera ley de Mendel, también llamada Ley de la Herencia Independiente de Caracteres o Ley de la Asociación Independiente. Según Mendel, hay rasgos heredados que se obtienen de forma independiente, sin relación con el fenotipo, lo cual no afecta al patrón de herencia de otros rasgos. Esta ley se cumple en los genes que no están ligados, es decir que se encuentran en diferentes cromosomas o que están en zonas muy separadas del mismo cromosoma.
Mendel, para concluir la tercera de las leyes de Mendel, realizó un cruce de plantas de chícharos que producían semillas amarillas y llanas, con chícharos que producían semillas verdes y con textura irregular. Estas eran homocigóticas para los dos caracteres de textura y color. Se concluía que la ley de uniformidad estaba presente, pues con la primera generación se pudo obtener semillas amarillas y lisas.
Sin embargo, al cruzar esta primera generación para obtener una segunda generación, se observan nuevos tipos de semillas con caracteres diversos pero relacionados con la generación parental, se obtuvieron semillas amarillas y lisas, amarillas y rugosas, verdes y lisas, y verdes y rugosas. Sigue leyendo aquí para aprender más acerca de Tercera Ley De Mendel.
SEMANA DEL 3 AL 7 DE ABRIL
SEMANA DEL 3 AL 7 DE ABRIL
Leyes De Mendel
 Las leyes de Mendel fueron desarrolladas por un científico genetista, considerado como el padre de la genética: Gregor Mendel. De allí su nombre. Este científico realizo experimentos que permitieron dilucidar elementos fundamentales de la herencia genética, como con un ejemplo de ley de Mendel, donde se explican los rasgos descendientes que se pueden predecir a través de las características de los progenitores de una especie, desde animales, plantas y hasta seres humanos.
Este científico fue quien acuñó algunos de los términos más conocidos de la genética, como son los términos “dominante” y “recesivo”, que son factores de la herencia presentes en las características y rasgos hereditarios en los organismos, todo esto a través de las tres Leyes de Mendel.
Las leyes de Mendel fueron desarrolladas por un científico genetista, considerado como el padre de la genética: Gregor Mendel. De allí su nombre. Este científico realizo experimentos que permitieron dilucidar elementos fundamentales de la herencia genética, como con un ejemplo de ley de Mendel, donde se explican los rasgos descendientes que se pueden predecir a través de las características de los progenitores de una especie, desde animales, plantas y hasta seres humanos.
Este científico fue quien acuñó algunos de los términos más conocidos de la genética, como son los términos “dominante” y “recesivo”, que son factores de la herencia presentes en las características y rasgos hereditarios en los organismos, todo esto a través de las tres Leyes de Mendel.
Fue decisivo el Ensayo Sobre Los Híbridos Vegetales que realizó en 1866, donde finalmente se formulaban las 3 Leyes De Mendel que fueron nombradas ante su apellido. Y que estaban compuestas por cruces inter-especies y experimentos que fueron llevados a un análisis estadístico. Sin embargo, estos estudios no fueron tomados en cuenta hasta mucho después de ser publicados, en el año 1900. A continuación te damos una introducción a las Leyes de Gregor Mendel:

Las leyes de Mendel fueron desarrolladas por un científico genetista, considerado como el padre de la genética: Gregor Mendel. De allí su nombre. Este científico realizo experimentos que permitieron dilucidar elementos fundamentales de la herencia genética, como con un ejemplo de ley de Mendel, donde se explican los rasgos descendientes que se pueden predecir a través de las características de los progenitores de una especie, desde animales, plantas y hasta seres humanos.
Este científico fue quien acuñó algunos de los términos más conocidos de la genética, como son los términos “dominante” y “recesivo”, que son factores de la herencia presentes en las características y rasgos hereditarios en los organismos, todo esto a través de las tres Leyes de Mendel.
Fue decisivo el Ensayo Sobre Los Híbridos Vegetales que realizó en 1866, donde finalmente se formulaban las 3 Leyes De Mendel que fueron nombradas ante su apellido. Y que estaban compuestas por cruces inter-especies y experimentos que fueron llevados a un análisis estadístico. Sin embargo, estos estudios no fueron tomados en cuenta hasta mucho después de ser publicados, en el año 1900. A continuación te damos una introducción a las Leyes de Gregor Mendel:
Fue decisivo el Ensayo Sobre Los Híbridos Vegetales que realizó en 1866, donde finalmente se formulaban las 3 Leyes De Mendel que fueron nombradas ante su apellido. Y que estaban compuestas por cruces inter-especies y experimentos que fueron llevados a un análisis estadístico. Sin embargo, estos estudios no fueron tomados en cuenta hasta mucho después de ser publicados, en el año 1900. A continuación te damos una introducción a las Leyes de Gregor Mendel:
Ley De Mendel
 La primera ley de Mendel, también llamada: Ley de la uniformidad de los híbridos de la primera generación, o simplemente Ley de la Uniformidad. Esta ley dicta que, al cruzar dos variedades de una especie de raza pura, cada uno de los híbridos de la primera generación tendrá caracteres determinados similares en su fenotipo. Esto se debe a que las razas puras tienen un gen dominante o un gen recesivo. El genotipo dominante será entonces el que determine la característica o características principales de la primera generación del cruce, pero al mismo tiempo, también serán similares fenotípicamente entre sí, es decir, entre cada individuo de la primera generación.
En el experimento realizado por Mendel para obtener la primera de las leyes de Mendel, utilizaba una especie de chícharos que producían semillas amarillas como gen dominante y otra que tenía un gen recesivo que producía semillas verdes, por lo tanto, el alelo que llamaremos “A” daba el color amarillo por encima del alelo “a” que producía el color verde. El producto del cruce eran plantas que producían semillas amarillas. Sigue leyendo aquí para aprender más sobre la Ley De Mendel.
La primera ley de Mendel, también llamada: Ley de la uniformidad de los híbridos de la primera generación, o simplemente Ley de la Uniformidad. Esta ley dicta que, al cruzar dos variedades de una especie de raza pura, cada uno de los híbridos de la primera generación tendrá caracteres determinados similares en su fenotipo. Esto se debe a que las razas puras tienen un gen dominante o un gen recesivo. El genotipo dominante será entonces el que determine la característica o características principales de la primera generación del cruce, pero al mismo tiempo, también serán similares fenotípicamente entre sí, es decir, entre cada individuo de la primera generación.
En el experimento realizado por Mendel para obtener la primera de las leyes de Mendel, utilizaba una especie de chícharos que producían semillas amarillas como gen dominante y otra que tenía un gen recesivo que producía semillas verdes, por lo tanto, el alelo que llamaremos “A” daba el color amarillo por encima del alelo “a” que producía el color verde. El producto del cruce eran plantas que producían semillas amarillas. Sigue leyendo aquí para aprender más sobre la Ley De Mendel.
La primera ley de Mendel, también llamada: Ley de la uniformidad de los híbridos de la primera generación, o simplemente Ley de la Uniformidad. Esta ley dicta que, al cruzar dos variedades de una especie de raza pura, cada uno de los híbridos de la primera generación tendrá caracteres determinados similares en su fenotipo. Esto se debe a que las razas puras tienen un gen dominante o un gen recesivo. El genotipo dominante será entonces el que determine la característica o características principales de la primera generación del cruce, pero al mismo tiempo, también serán similares fenotípicamente entre sí, es decir, entre cada individuo de la primera generación.
En el experimento realizado por Mendel para obtener la primera de las leyes de Mendel, utilizaba una especie de chícharos que producían semillas amarillas como gen dominante y otra que tenía un gen recesivo que producía semillas verdes, por lo tanto, el alelo que llamaremos “A” daba el color amarillo por encima del alelo “a” que producía el color verde. El producto del cruce eran plantas que producían semillas amarillas. Sigue leyendo aquí para aprender más sobre la Ley De Mendel.
Segunda Ley De Mendel
La segunda ley de Mendel, también conocida como la Ley de la Segregación, Ley de la Separación Equitativa, o hasta Ley de Disyunción de los Alelos. Esta dictamina que para que exista la reproducción de dos individuos de una especie, primero debe existir la separación del alelo de cada uno de los pares para que de esta manera se transfiera la información genética al hijo. Un alelo es, la variante genética que permite determinar un rasgo o carácter. Existen entonces, alelos dominantes y alelos recesivos.
 Por esto, es que la segunda de las leyes de Mendel se la llama como de segregación o separación, ya que cada padre, aporta un alelo que se separa de cada uno, para formar un individuo en una nueva generación. Sigue leyendo aquí para aprender más sobre la Segunda Ley De Mendel. Mendel, en su experimento, obtuvo solo semillas amarillas en la primera generación, pero en la segunda generación, los alelos se separaron para formar nuevas semillas verdes en menor proporción que las amarillas, pero aun así existentes. Esta sería la proporción:
Por esto, es que la segunda de las leyes de Mendel se la llama como de segregación o separación, ya que cada padre, aporta un alelo que se separa de cada uno, para formar un individuo en una nueva generación. Sigue leyendo aquí para aprender más sobre la Segunda Ley De Mendel. Mendel, en su experimento, obtuvo solo semillas amarillas en la primera generación, pero en la segunda generación, los alelos se separaron para formar nuevas semillas verdes en menor proporción que las amarillas, pero aun así existentes. Esta sería la proporción:
La segunda ley de Mendel, también conocida como la Ley de la Segregación, Ley de la Separación Equitativa, o hasta Ley de Disyunción de los Alelos. Esta dictamina que para que exista la reproducción de dos individuos de una especie, primero debe existir la separación del alelo de cada uno de los pares para que de esta manera se transfiera la información genética al hijo. Un alelo es, la variante genética que permite determinar un rasgo o carácter. Existen entonces, alelos dominantes y alelos recesivos.
Por esto, es que la segunda de las leyes de Mendel se la llama como de segregación o separación, ya que cada padre, aporta un alelo que se separa de cada uno, para formar un individuo en una nueva generación. Sigue leyendo aquí para aprender más sobre la Segunda Ley De Mendel. Mendel, en su experimento, obtuvo solo semillas amarillas en la primera generación, pero en la segunda generación, los alelos se separaron para formar nuevas semillas verdes en menor proporción que las amarillas, pero aun así existentes. Esta sería la proporción:
Tercera Ley De Mendel
La tercera ley de Mendel, también llamada Ley de la Herencia Independiente de Caracteres o Ley de la Asociación Independiente. Según Mendel, hay rasgos heredados que se obtienen de forma independiente, sin relación con el fenotipo, lo cual no afecta al patrón de herencia de otros rasgos. Esta ley se cumple en los genes que no están ligados, es decir que se encuentran en diferentes cromosomas o que están en zonas muy separadas del mismo cromosoma.
Mendel, para concluir la tercera de las leyes de Mendel, realizó un cruce de plantas de chícharos que producían semillas amarillas y llanas, con chícharos que producían semillas verdes y con textura irregular. Estas eran homocigóticas para los dos caracteres de textura y color. Se concluía que la ley de uniformidad estaba presente, pues con la primera generación se pudo obtener semillas amarillas y lisas.
Sin embargo, al cruzar esta primera generación para obtener una segunda generación, se observan nuevos tipos de semillas con caracteres diversos pero relacionados con la generación parental, se obtuvieron semillas amarillas y lisas, amarillas y rugosas, verdes y lisas, y verdes y rugosas. Sigue leyendo aquí para aprender más acerca de Tercera Ley De Mendel.
La tercera ley de Mendel, también llamada Ley de la Herencia Independiente de Caracteres o Ley de la Asociación Independiente. Según Mendel, hay rasgos heredados que se obtienen de forma independiente, sin relación con el fenotipo, lo cual no afecta al patrón de herencia de otros rasgos. Esta ley se cumple en los genes que no están ligados, es decir que se encuentran en diferentes cromosomas o que están en zonas muy separadas del mismo cromosoma.
Mendel, para concluir la tercera de las leyes de Mendel, realizó un cruce de plantas de chícharos que producían semillas amarillas y llanas, con chícharos que producían semillas verdes y con textura irregular. Estas eran homocigóticas para los dos caracteres de textura y color. Se concluía que la ley de uniformidad estaba presente, pues con la primera generación se pudo obtener semillas amarillas y lisas.
Sin embargo, al cruzar esta primera generación para obtener una segunda generación, se observan nuevos tipos de semillas con caracteres diversos pero relacionados con la generación parental, se obtuvieron semillas amarillas y lisas, amarillas y rugosas, verdes y lisas, y verdes y rugosas. Sigue leyendo aquí para aprender más acerca de Tercera Ley De Mendel.
Leyes De Mendel Resumidas
Para terminar con las Leyes De Mendel Resumidas, se puede decir que, la primera Ley de Mendel dice que si se cruzan dos padres de raza pura con diferentes rasgos, la primera generación tendrá similitudes entre sí y guardará un carácter del padre con el alelo dominante. La segunda ley dice que, los factores genéticos se separan de cada uno de los padres en alelos individuales que se juntarán para procrear una descendencia con las características de la primera generación, pero en la segunda generación, se manifiestan nuevos rasgos genéticos observados en los padres pero unidos de manera aleatoria en la descendencia de la primera generación. Y la tercera ley de Mendel dice que, además existen rasgos generados de forma independiente, a través de cromosomas alejados que no intervienen entre sí, y al igual que en la segunda ley, esta tercera de las leyes de Mendel se manifiesta con más claridad en la segunda generación de individuos.
DEL 27 AL 31 DE MARZO
PRIMER PERIODO
LAS MUTACIONES
La Mutación es cualquier alteración o variación en el código genético; es decir, una alteración de los genes de los cromosomas. Es posible que una mutación ocurra mientras se realiza la meiosis.
Esta variación puede producirse en las células somáticas o en las células sexuales (gametos). Si las mutaciones se presentan en el ADN de los gametos, pueden transmitirse de una generación a otra. Por el contrario, si se produce en las células somáticas, no se heredará, pero puede propagarse asexualmente, lo cual sucede en las plantas (por ejemplo, las que se reproducen por estacas).
Las mutaciones pueden ocurrir de manera espontánea o inducida por algunos agentes llamados mutágenos, se clasifican en externos e internos. Los agentes externos pueden ser las radiaciones ultravioleta, los rayos X, los cambios de temperatura, determinadas sustancias químicas, entre otros. Los agentes internos son los cambios accidentales del código del ADN o la ausencia de sectores del gen o del cromosoma
Pocas ideas han cambiado tan profundamente nuestra visión de la naturaleza como la misma idea de cambio que implica la evolución de los seres vivos. Los organismos biológicos se agrupan en unidades naturales de reproducción que denominamos especies. Las especies que ahora pueblan la Tierra proceden de otras especies distintas que existieron en el pasado, a través de un proceso de descendencia con modificación. La evolución biológica es el proceso histórico de transformación de unas especies en otras especies descendientes, y su reverso es la extinción de la gran mayoría de las especies que han existido. Una de las ideas más románticas contenidas en la evolución de la vida es que dos organismos vivos cualesquiera, por diferentes que sean, comparten un antecesor común en algún momento del pasado. Nosotros y cualquier chimpancé actual compartimos un antepasado hace algo así como 5 millones años. También tenemos un antecesor común con cualquiera de las bacterias hoy existentes, aunque el tiempo al antecesor se remonte en este caso a más de 3000 millones de años.

La idea de evolución por modificación y derivación de nuevas especies implica la existencia de antepasados comunes para cualquier par de especies. Hay un antepasado común del hombre y el chimpancé, y del hombre y las bacterias.
La evolución es el gran principio unificador de la Biología, sin ella no es posible entender ni las propiedades distintivas de los organismos, sus adaptaciones; ni las relaciones de mayor o menor proximidad que existen entre las distintas especies. La teoría evolutiva se relaciona con el resto de la biología de forma análoga a como el estudio de la historia se relaciona con las ciencias sociales. La famosa frase del genético evolucionista Theodosius Dobzhansky que abre este tema, no es más que una aplicación particular del principio más general que afirma que el presente de un proceso temporal no puede entenderse sin una perspectiva histórica.
Para terminar con las Leyes De Mendel Resumidas, se puede decir que, la primera Ley de Mendel dice que si se cruzan dos padres de raza pura con diferentes rasgos, la primera generación tendrá similitudes entre sí y guardará un carácter del padre con el alelo dominante. La segunda ley dice que, los factores genéticos se separan de cada uno de los padres en alelos individuales que se juntarán para procrear una descendencia con las características de la primera generación, pero en la segunda generación, se manifiestan nuevos rasgos genéticos observados en los padres pero unidos de manera aleatoria en la descendencia de la primera generación. Y la tercera ley de Mendel dice que, además existen rasgos generados de forma independiente, a través de cromosomas alejados que no intervienen entre sí, y al igual que en la segunda ley, esta tercera de las leyes de Mendel se manifiesta con más claridad en la segunda generación de individuos.
PRIMER PERIODO
LAS MUTACIONES
La Mutación es cualquier alteración o variación en el código genético; es decir, una alteración de los genes de los cromosomas. Es posible que una mutación ocurra mientras se realiza la meiosis.
Esta variación puede producirse en las células somáticas o en las células sexuales (gametos). Si las mutaciones se presentan en el ADN de los gametos, pueden transmitirse de una generación a otra. Por el contrario, si se produce en las células somáticas, no se heredará, pero puede propagarse asexualmente, lo cual sucede en las plantas (por ejemplo, las que se reproducen por estacas).
Las mutaciones pueden ocurrir de manera espontánea o inducida por algunos agentes llamados mutágenos, se clasifican en externos e internos. Los agentes externos pueden ser las radiaciones ultravioleta, los rayos X, los cambios de temperatura, determinadas sustancias químicas, entre otros. Los agentes internos son los cambios accidentales del código del ADN o la ausencia de sectores del gen o del cromosoma
Pocas ideas han cambiado tan profundamente nuestra visión de la naturaleza como la misma idea de cambio que implica la evolución de los seres vivos. Los organismos biológicos se agrupan en unidades naturales de reproducción que denominamos especies. Las especies que ahora pueblan la Tierra proceden de otras especies distintas que existieron en el pasado, a través de un proceso de descendencia con modificación. La evolución biológica es el proceso histórico de transformación de unas especies en otras especies descendientes, y su reverso es la extinción de la gran mayoría de las especies que han existido. Una de las ideas más románticas contenidas en la evolución de la vida es que dos organismos vivos cualesquiera, por diferentes que sean, comparten un antecesor común en algún momento del pasado. Nosotros y cualquier chimpancé actual compartimos un antepasado hace algo así como 5 millones años. También tenemos un antecesor común con cualquiera de las bacterias hoy existentes, aunque el tiempo al antecesor se remonte en este caso a más de 3000 millones de años.
La idea de evolución por modificación y derivación de nuevas especies implica la existencia de antepasados comunes para cualquier par de especies. Hay un antepasado común del hombre y el chimpancé, y del hombre y las bacterias.
La evolución es el gran principio unificador de la Biología, sin ella no es posible entender ni las propiedades distintivas de los organismos, sus adaptaciones; ni las relaciones de mayor o menor proximidad que existen entre las distintas especies. La teoría evolutiva se relaciona con el resto de la biología de forma análoga a como el estudio de la historia se relaciona con las ciencias sociales. La famosa frase del genético evolucionista Theodosius Dobzhansky que abre este tema, no es más que una aplicación particular del principio más general que afirma que el presente de un proceso temporal no puede entenderse sin una perspectiva histórica.
La revolución darwiniana
Aunque la idea de la evolución tenía precedentes, no fue hasta 1859, con la aparición de la obra El origen de las especies del naturalista británico Charles Darwin, que la idea de la evolución se estableció definitivamente. Darwin recopiló e interpretó un gran número de observaciones y experimentos de muy diversas disciplinas de investigación y los presentó como un argumento irrefutable en favor del hecho de la evolución. Pero Darwin suministró además un mecanismo para explicar las adaptaciones complejas y características de los seres vivos: la selección natural. ¿Qué significó la teoría de la evolución y de la selección natural en el contexto de la biología del siglo XIX? En 1802 el teólogo W. Paley publica la obra Teología natural, en donde arguye que el diseño funcional de los organismos evidenciaba la existencia de un creador omnisapiente. Según él, el ojo humano, con su delicado diseño, constituía una prueba concluyente de la existencia de Dios. Para los naturalistas que querían explicar los fenómenos biológicos por procesos naturales, explicar la adaptación, la maravillosa adecuación de los organismos a su ambiente, constituía el problema fundamental.

 El gran reto de Darwin era explicar las complejas adaptaciones de los organismos vivos, como el diseño funcional de un ojo, por mecanismos naturales. La solución de Darwin fue proponer el mecanismo de la selección natural
El gran reto de Darwin era explicar las complejas adaptaciones de los organismos vivos, como el diseño funcional de un ojo, por mecanismos naturales. La solución de Darwin fue proponer el mecanismo de la selección natural
El argumento del diseño de Paley tenía una gran influencia en los naturalistas del XIX, a pesar de que esta visión intervencionista violaba flagrantemente el concepto de naturaleza que se había establecido con el desarrollo de la física en los siglos XVI y XVII. Los fenómenos del Universo, según esta nueva concepción, eran explicables por procesos naturales. La naturaleza, per se, era un objeto lícito para preguntar y contestar científicamente. Con el Origen de Darwin se introduce esta revolución en la Biología. Lo verdaderamente revolucionario en Darwin fue el proponer un mecanismo natural para explicar la génesis, diversidad y adaptación de los organismos.
 El naturalista británico Charles Darwin (1809-1882) introdujo en su libro El origen de las especies (1859) dos ideas revolucionarias: la evolución biológica y la selección natural
El naturalista británico Charles Darwin (1809-1882) introdujo en su libro El origen de las especies (1859) dos ideas revolucionarias: la evolución biológica y la selección natural
El pensamiento poblacional
Hay grandeza en esta concepción de la vida,... que mientras este planeta ha ido girando según la constante ley de la gravitación, se han desarrollado y se están desarrollando, a partir de un comienzo tan sencillo, infinidad de formas cada vez más bellas y maravillosas
Charles Darwin
Para imponer su teoría de la evolución y de la selección natural, Darwin tuvo que introducir una nueva forma de entender la variación en la naturaleza, el pensamiento poblacional. En el tiempo de Darwin las especies se consideraban entidades fijas e inmutables; representaban a un tipo platónico, la idea perfecta de la mente de su creador. Las diferencias en la forma, en la conducta, o en la fisiología de los organismos de una especie no eran más que imperfecciones, errores en la materialización de la idea de la especie. En contraste con esta visión esencialista dominante, la variación individual, lejos de ser trivial, es para Darwin la piedra angular de la evolución. La variación en el seno de las especies o poblaciones es lo único real, es la materia prima de la evolución, a partir de la que se va a crear toda la diversidad biológica. Son las diferencias existentes entre los organismos de una especie las que, al magnificarse en el espacio y en el tiempo, producirán nuevas poblaciones, nuevas especies, y por extensión, toda la diversidad biológica.
 Bajo la visión darwiniana, la variación es la única realidad de las especies. No hay un color de piel en la especie humana ideal o arquetípico. Cada individuo con su variación característica es un elemento esencial de nuestra especie
Bajo la visión darwiniana, la variación es la única realidad de las especies. No hay un color de piel en la especie humana ideal o arquetípico. Cada individuo con su variación característica es un elemento esencial de nuestra especie
Aunque la idea de la evolución tenía precedentes, no fue hasta 1859, con la aparición de la obra El origen de las especies del naturalista británico Charles Darwin, que la idea de la evolución se estableció definitivamente. Darwin recopiló e interpretó un gran número de observaciones y experimentos de muy diversas disciplinas de investigación y los presentó como un argumento irrefutable en favor del hecho de la evolución. Pero Darwin suministró además un mecanismo para explicar las adaptaciones complejas y características de los seres vivos: la selección natural. ¿Qué significó la teoría de la evolución y de la selección natural en el contexto de la biología del siglo XIX? En 1802 el teólogo W. Paley publica la obra Teología natural, en donde arguye que el diseño funcional de los organismos evidenciaba la existencia de un creador omnisapiente. Según él, el ojo humano, con su delicado diseño, constituía una prueba concluyente de la existencia de Dios. Para los naturalistas que querían explicar los fenómenos biológicos por procesos naturales, explicar la adaptación, la maravillosa adecuación de los organismos a su ambiente, constituía el problema fundamental.
El gran reto de Darwin era explicar las complejas adaptaciones de los organismos vivos, como el diseño funcional de un ojo, por mecanismos naturales. La solución de Darwin fue proponer el mecanismo de la selección natural
El argumento del diseño de Paley tenía una gran influencia en los naturalistas del XIX, a pesar de que esta visión intervencionista violaba flagrantemente el concepto de naturaleza que se había establecido con el desarrollo de la física en los siglos XVI y XVII. Los fenómenos del Universo, según esta nueva concepción, eran explicables por procesos naturales. La naturaleza, per se, era un objeto lícito para preguntar y contestar científicamente. Con el Origen de Darwin se introduce esta revolución en la Biología. Lo verdaderamente revolucionario en Darwin fue el proponer un mecanismo natural para explicar la génesis, diversidad y adaptación de los organismos.
El naturalista británico Charles Darwin (1809-1882) introdujo en su libro El origen de las especies (1859) dos ideas revolucionarias: la evolución biológica y la selección natural
El pensamiento poblacional
Hay grandeza en esta concepción de la vida,... que mientras este planeta ha ido girando según la constante ley de la gravitación, se han desarrollado y se están desarrollando, a partir de un comienzo tan sencillo, infinidad de formas cada vez más bellas y maravillosas
Charles Darwin
Para imponer su teoría de la evolución y de la selección natural, Darwin tuvo que introducir una nueva forma de entender la variación en la naturaleza, el pensamiento poblacional. En el tiempo de Darwin las especies se consideraban entidades fijas e inmutables; representaban a un tipo platónico, la idea perfecta de la mente de su creador. Las diferencias en la forma, en la conducta, o en la fisiología de los organismos de una especie no eran más que imperfecciones, errores en la materialización de la idea de la especie. En contraste con esta visión esencialista dominante, la variación individual, lejos de ser trivial, es para Darwin la piedra angular de la evolución. La variación en el seno de las especies o poblaciones es lo único real, es la materia prima de la evolución, a partir de la que se va a crear toda la diversidad biológica. Son las diferencias existentes entre los organismos de una especie las que, al magnificarse en el espacio y en el tiempo, producirán nuevas poblaciones, nuevas especies, y por extensión, toda la diversidad biológica.
Bajo la visión darwiniana, la variación es la única realidad de las especies. No hay un color de piel en la especie humana ideal o arquetípico. Cada individuo con su variación característica es un elemento esencial de nuestra especie
El estudio de la evolución
Los estudios y afirmaciones acerca de la evolución generalmente se refieren a uno de dos aspectos distintos: (1) las investigaciones acerca del hecho de la evolución y (2) las que se refieren al mecanismo de la evolución. Las primeras abarcan las disciplinas biológicas, tales como la paleontología, la clasificación, la biología comparada, la biología de poblaciones,... que muestran de manera inequívoca el hecho de la evolución. Las segundas, las afirmaciones acerca del mecanismo de la evolución, son el objeto principal de estudio de la genética de poblaciones, y nos informan de los factores, fuerzas o procesos que producen el cambio evolutivo, es decir, los mecanismos naturales que causan la descendencia con modificación. Una analogía cotidiana que ilustra esta distinción es la del tiempo meteorológico. Las precipitaciones, los vientos, las gotas frías, los tifones, son las evidencias que constituyen las afirmaciones de hecho del tiempo atmosférico. Ahora bien, si queremos explicar el porqué se dan los diferentes fenómenos meteorológicos, entonces nos tenemos que introducir en el ámbito de las afirmaciones del proceso o de los mecanismos meteorológicos. Debemos proponer los factores, tales como las diferencias de temperatura entre distintas masas de aire, que producen los fenómenos meteorológicos.
La evidencia de la evolución
La evolución que se da en una escala reducida, en el interior de una especie y en el intervalo de unas pocas generaciones, se denomina microevolución. La macroevolución es la evolución a gran escala, y abarca periodos considerables de tiempo, y grandes procesos de transformación; en el caso más extremo comprendería toda la evolución de la vida. Se pueden efectuar experimentos y/o observaciones de poblaciones de especies actuales a pequeña escala y obtener evidencia directa de evolución. Hay muchos ejemplos en los que se detecta la evolución en acción, como el clásico caso del melanismo industrial que se comentará más adelante. La selección artificial efectuada por el hombre en el perro o el caballo son también claros ejemplos que muestran el potencial de modificación de una especie. Por su propia dimensión temporal, no podemos demostrar la macroevolución directamente, exceptuando los casos de creación de nuevas especies de plantas mediante cruzamiento de especies distintas por el hombre. Aunque la evidencia evolutiva que suministra el registro fósil, la biología comparada, y la biología molecular es indirecta, no por ello es menos concluyente a la hora de demostrar la comunidad de origen de todos los organismos.

Las diferentes razas de perro que el hombre ha obtenido por selección artificial ilustran el potencial de cambio que tienen las especies.
El registro fósil
Los sedimentos que se han ido acumulando sobre la corteza de la tierra durante su historia geológica dejan una huella inestimable, generalmente en forma de huesos o esqueletos duros petrificados, de organismos muertos en el pasado: son los fósiles. El registro fósil es una ventana maravillosa a la historia de la vida. Si no existiera no podríamos inventarlo. Sin él, el vacío acerca de la evolución de la vida sobre la tierra sería insustituible. Podríamos especular, teorizar infinitamente, pero ¿quién podría haber imaginado que la Tierra estuvo dominada durante 150 millones de años por unos reptiles inmensos y fantásticos, los dinosaurios, que desaparecieron en un instante relativo de tiempo, si no hubieran existido fósiles de dinosaurios que nos lo contasen? La desintegración de los elementos químicos radiactivos que hay en las rocas ha permitido estimar que la Tierra se originó alrededor de hace 4600 millones de años. La Tierra, que era una esfera caliente, se enfría gradualmente, iniciándose un periodo de evolución química que culminará con la formación de las primeras células. En Australia y África se han encontrado sedimentos retenidos y fijados por bacterias de hace 3600 millones de años, lo que hace que esta fecha sea una estima mínima de la edad de inicio de la evolución biológica. La magnitud del tiempo en el que ha transcurrido la evolución se escapa completamente a nuestra comprensión, no podemos siquiera imaginar, limitados a la minúscula escala de nuestro tiempo vital, el potencial de transformación que suponen 3600 millones de años de evolución.
El tiempo geológico
El tiempo geológico se ha dividido en una serie de etapas jerárquicas, las eras, los períodos, y las épocas, que no siguen una cronología lineal, sino que es una crónica de los momentos claves de la historia de la vida. Las transiciones entre las cuatro eras, la Precámbrica, la Paleozoica, la Mesozoica y la Cenozoica representan grandes cambios en las fauna y flora de toda la Tierra. En el primer período de la era Paleozoica, el Cámbrico, hace 570 millones de años, aparecen de golpe en el registro fósil los animales pluricelulares que tienen partes duras, como las conchas, y exoesqueletos,... El final del Paleozoico coincide con la mayor extinción habida en la Tierra, en la que desaparecieron el 96% de las especies. Al final del Mesozoico, en la transición entre el período Cretácico y Terciario, se da la conocida extinción de los dinosaurios, junto a un 70% de las especies existentes.
Cronología de siete momentos estelares de la evolución
¿Qué nos enseña el registro fósil sobre la historia de la vida sobre la Tierra? Esta es una lista de los acontecimientos más importantes
-
Origen de la célula procariota 3600 M (M=Millones de años)
-
Origen de la célula eucariota 1400 M
-
Origen de la fauna de animales pluricelulares 650 M
-
Fauna de la explosión cámbrica 570 M
-
Origen de los vertebrados terrestres 360 M
-
Extinción de los dinosaurios. La antorcha pasa de los dinosaurios a los mamíferos 65 M
-
Origen de Homo sapiens 0,1 M.
Si toda la historia de la Tierra la comprimiésemos en una hora, a los 20 minutos aparecerían las bacterias, a los 55 los dinosaurios, los antropoides aparecen a 40 segundos antes del final, y los humanos al cumplirse la hora.
Hay una coherencia entre el registro fósil y el origen evolutivo de las especies y sus grupos correspondientes. Si la vida se debiera a la creación espontánea e independiente de especies, no tendríamos porqué esperar que el hombre surgiera con los mamíferos, podríamos haber surgido en cualquier momento, cuando aparecieron los peces, o los dinosaurios, o antes de que surgieran los primates.
La evolución no tiene dirección definida
La historia de la vida es una historia de extinciones y muerte,... con unos pocos supervivientes. El 99,9% de las especies que han existido alguna vez están hoy extintas. Grupos enteros de organismos, como los dinosaurios (exceptúando las aves), los trilobites, los ammonoideos, se han extinguido para siempre sin dejar descendiente alguno.
Como señala el reconocido paleontólogo S. Gould, el registro fósil no es un relato convencional que conduce a los diferentes linajes a más excelencia, más complejidad, más diversidad. La historia de la vida no muestra un rumbo definido, no tiene dirección ni sentido. La evolución es una narración de eliminación masiva seguida de diferenciación en el interior de unos cuantos supervivientes. Es a priori imposible determinar la dirección de la evolución porque la importancia de los acontecimientos concretos, contingentes, como la extinción o no de un grupo de organismos en el caso de una extinción en masa, o la posesión o no de una variante adaptativa adecuada cuando ésta es requerida, son los verdaderos agentes de la historia.
Las extinciones en masa
Hay dos regímenes de extinciones: la extinción normal, que afecta a las especies que dejan de seguir a su ambiente en su lucha diaria por adaptarse, y la extinción en masa, que son más rápidas y devastadoras en cuanto a su magnitud. Al menos han ocurrido cinco extinciones en masa, y han dejado muchos huecos ecológicos que han permitido que sean ocupados por los descendientes de las especies supervivientes. Esta ocupación del bioespacio disponible suele ir acompañada de una rápida y extensa diversificación morfológica que se denomina radiación adaptativa. No se sabe con certeza la causa de las extinciones masivas, aunque las causas físicas como el impacto de asteroides o el cambio climático parecen más probables que las biológicas. Según una hipótesis reciente, hay ciclos de extinción masiva aproximadamente cada 26 millones de años, y el impacto periódico de lluvias de cometas sobre la Tierra podría explicar dicho ciclo.
La fauna de Burguess Shale
Si tuviéramos la oportunidad de volver a un momento del pasado, el período previo al Cámbrico sería sin duda uno de los más atractivos. En el Cámbrico, hace 570 M, se produce la gran explosión en el registro fósil de los primeros animales pluricelulares con partes duras. Darwin se preguntaba por qué estos primeros animales eran ya anatómicamente complejos y sin precursores aparentes. La respuesta está en el yacimiento de Burguess Shale, situado en las montañas rocosas canadienses. Debido a condiciones muy especiales de conservación, aquí se encuentra la única fauna de cuerpo blando (sin estructuras duras) que existe de un tiempo inmediatamente anterior a la explosión cámbrica.
La gran sorpresa de Burguess Shale es que las pocas especies analizadas contienen una disparidad de diseños anatómicos que exceden, con mucho, la gama moderna que hay en todo el mundo. En ninguna fauna posterior se repite la riqueza anatómica de este yacimiento. De los 120 géneros analizados, 20 son diseños de artrópodos únicos, y además de estar representados los 4 grupos de animales que hoy existen, hay 8 diseños que no encajan en ningún grupo animal conocido. La época de Burguess Shale parece ser que fue un tiempo asombroso de experimentación, una era de gran flexibilidad evolutiva, que fue seguido de una gran extinción.
Opabinia: una vida extraña de un remoto pasado
Opabinia fue el fósil de Burguess Shale que reveló la caja de sorpresas que había en este yacimiento. Su diseño singular no pertenece a ningún grupo animal conocido. ¡Tiene 5 ojos!, trompa flexible formada por un tubo cilíndrico estriado, un tubo digestivo en forma de U, un tronco de 15 segmentos, con lóbulos delgados laterales. Los 3 segmentos últimos forman una cola con tres pares de hojas delgadas y lobuladas dirigidas hacia afuera.

Opabinia fue el primer fósil del yacimiento de Burguess Shale que demostró la singularidad morfológica de las primeras formas pluricelulares. Tiene 5 ojos, trompa flexible, tubo digestivo en forma de U, un tronco de 15 segmentos, los 3 últimos forman una cola.
Los dinosaurios
Durante el periodo Triásico surgen los dinosaurios de una línea de los reptiles e inician una dinastía sobre el medio terrestre que habría de durar 150 M. Se han identificado más de 350 especies de dinosaurios, y se cree que esta es una estima muy inferior a la de las especies que realmente existieron. Entre los dinosaurios están los animales más grandes que alguna vez hayan vivido sobre la Tierra. El Seismosaurus, el mayor herbívoro conocido, tenía unas 40 metros de longitud. El Tyrannosaurus rex, uno de los mayores carnívoros, tenía 12 m de longitud. Los dinosaurios se dividen en dos grandes órdenes, los de pelvis de lagarto (Saurisquios), que comprenden tanto especies carnívoras como herbívoras, y los de pelvis de ave (Ornistiquios), cuyas especies eran todas herbívoras. Los dinosaurios se extinguieron casi en su totalidad, exceptuando una línea menor que dio lugar a las aves, hace 65 M. Junto a ellos, desaparecieron en un breve lapso de tiempo el 75% de las especies existentes. Se ha propuesto que el impacto de un asteroide sobre la superficie terrestre podría ser la causa de la extinción en masa.
 ¿Quién podría haber imaginado que la Tierra estuvo dominada durante 150 millones de años por unos reptiles inmensos y fantásticos, los dinosaurios, que desaparecieron en un instante relativo de tiempo, si no hubieran existido fósiles de dinosaurios que nos lo contasen?
¿Quién podría haber imaginado que la Tierra estuvo dominada durante 150 millones de años por unos reptiles inmensos y fantásticos, los dinosaurios, que desaparecieron en un instante relativo de tiempo, si no hubieran existido fósiles de dinosaurios que nos lo contasen?
Biología comparada: homología y analogía
Cuando uno observa similitudes entre especies, se pueden distinguir entre dos tipos de semejanzas, la analogía y la homología. El ala de un ave y el de una mosca forman una extensión plana y tienen un movimiento de aleteo similar; los peces, los delfines, o los pingüinos tienen una sección transversal aplanada que les permite desplazarse por el agua. Estas semejanzas, llamadas analogías, son más bien superficiales y se deben a que estos organismos están sometidos a las mismas restricciones funcionales o adaptativas, y no son debidas a que posean un antepasado común reciente.
En contraste con la analogía, una homología es la similitud que hay entre caracteres de distintas especies debido a que tienen un origen común, y no a la acción directa de una presión funcional. Por ejemplo, todos los tetrápodos (animales vertebrados terrestres con cuatro extremidades) tienen la extremidad de cinco dedos, y ésta se encuentra tanto en las alas de la aves y de murciélagos como en la mano del ser humano, a pesar que estas extremidades representan unos papeles funcionales muy distintos. La razón de esta estructura común es que todos los tetrápodos conservamos la misma estructura básica de la especie ancestral original.

Homología: todos los tetrápodos tienen una extremidad con cinco dedos, aunque tengan diferentes funciones.
La homología es la base de la clasificación
La clasificación se basa en la comparación de los caracteres de las especies, y los caracteres homólogos son los elementos claves para establecer una clasificación evolutiva. Si las especies proceden de otras especies por evolución, y además no varían tan rápidamente como para perder toda su herencia histórica, se esperaría que los distintos seres vivos compartieran una serie de caracteres homólogos. El análisis de los diferentes caracteres fenotípicos, como la morfología, la conducta, los cromosomas, la anatomía externa e interna, el desarrollo embrionario, el metabolismo, la variación genética y proteica muestran que las especies presentan semejanzas homológas en todos los niveles del fenotipo. Cuanto más próxima sean la especies, mayor será el grado de semejanza, y lo contrario también es cierto, cuanto más alejada estén menos semejanzas encontraremos. Así, las diferencias que hoy vemos entre las especies se deben a las nuevas variaciones que han adquirido desde su separación del antepasado común. Las similitudes que atribuimos a las homologías no podrían explicarse si las especies se originasen independientemente unas de otras.
Órganos vestigiales
Un caso especialmente significativo de homología es la de los órganos vestigiales o residuales. La pelvis de los tetrápodos es una estructura ósea cuya función es articular las extremidades posteriores. Las ballenas son tetrápodos cuyos antepasados mamíferos abandonaron la tierra para habitar en el mar. En este nuevo medio las ballenas perdieron sus extremidades inferiores, pero aún conservan como huella acusatoria de su pasado tetrápodo, la serie completa de los huesos de la pelvis. De forma similar, las serpientes presentan vestigios de la extremidad posterior que portaban sus antepasados.

La reducida pelvis de los cetáceos es un órgano vestigial que prueba su origen tetrápodo.
Biología molecular y homología
La biología molecular ha suministrado la evidencia más universal de homología. Todos los organismos vivos compartimos el mismo material hereditario, el DNA, una molécula helicoidal cuya información se encuentra codificada en 4 letras o nucleótidos distintos. Igualmente, el código genético es prácticamente Universal, todos los organismos comparten el mismo diccionario que da el significado a la secuencia de DNA. Ambos ejemplos son pruebas muy robustas de la relación íntima que existe entre lo viviente.

El DNA es una molécula helicoidal que tiene información genética codificada a partir de cuatro letras o nucleótidos distintos.
El árbol de la vida: filogenia
Si la historia de la vida es cambio y ramificación por descendencia, entonces su representación sería la de un árbol o filogenia, en la que el tronco y las ramas internas se corresponderían a los antepasados de las especies actuales y los extremos de las ramas externas serían las especies actuales. ¿Como se establece una filogenia? Ordenando las especies actuales atendiendo a la similaridad morfológica y/o genética de sus caracteres homólogos.
El sistema linneano
El botánico sueco Carolus Linnaeus (1707-78) ideó el sistema de clasificación que se utiliza, con algunas modificaciones, hoy en día. Hay siete niveles inclusivos de clasificación, que son, de menor a mayor, la especie, el género, la familia, el orden, la clase, el tipo o Phylum y el reino. El nombre científico de cada especie tiene dos partes, el león, por ejemplo, se denomina Panthera leo. La primera parte se refiere al género y la segunda a la especie. Consideremos un ejemplo de como se agrupan las especies actuales en las distintas categorías linneanas. El león, la pantera, el tigre, pertenecen al género Panthera, que junto al género Felix (el gato doméstico) y otros se agrupan en la familia de los felinos. Los felinos, con los cánidos y úrsidos, constituyen el orden de los carnívoros. Primates, roedores, carnívoros, ... se reúnen en la clase mamíferos. Estos organismos comparten características como el amamantar a sus crías con leche, gestarlas en el útero mediante un órgano complejo, la placenta. Su piel está protegida por pelaje o pelos. Mamíferos, aves, reptiles, anfibios, y peces se reúnen en un solo tipo o phylum, porque todos tienen columna vertebral, un máximo de cuatro miembros, y sangre roja con hemoglobina, son los cordados. Los insectos, las arañas, los crustáceos, y los ciempiés, se clasifican en otro tipo, los artrópodos. Las almejas, caracoles y calamares se agrupan en los moluscos, y así sucesivamente. El tipo cordado, artrópodo, molusco y otros forman el reino Animal. El árbol de la vida se ordena siguiendo divisiones que van de características generales a aspectos más específicos.
 El botánico sueco Carolus Linnaeus (1707-78) creó el sistema de clasificación que se utiliza hoy en día.
El botánico sueco Carolus Linnaeus (1707-78) creó el sistema de clasificación que se utiliza hoy en día.
Significado de los niveles jerárquicos de clasificación
¿Por qué se ordena la diversidad biológica en diferentes niveles jerárquicos? Los rasgos de las divisiones más generales corresponderían a adaptaciones básicas o principales que surgieron en los momentos iniciales de la evolución de los especies progenitoras de estos grupos. Por ejemplo, hay cinco grandes reinos, las moneras, los protistas, los hongos, las plantas y los animales, que se corresponden con las cinco diferenciaciones principales de la vida sobre la Tierra. El Phylum o tipo es la siguiente unidad básica de diferenciación entre reinos, y podrían entenderse como proyectos básicos fundamentales de anatomía. En el reino Animal estos planos fundamentales serían las esponjas, los anélidos (gusanos), los artrópodos (insectos, crustáceos,...), los cordados, .... A partir de estas adaptaciones principales surgen posteriormente nuevas variaciones o subadaptaciones diversas, que por tener un menor tiempo de evolución, tienen un rango clasificatorio menor.
La clasificación de la diversidad biológica es una prueba de la evolución
El hecho que la diversidad de la vida esté jerarquizada es un fuerte argumento en favor de la evolución. El que las especies compartan estructuras anatómicas y adaptaciones básicas puede explicarse fácilmente si suponemos que las especies actuales compartieron antepasados, pero no esperaríamos dicho patrón si las especies hubieran sido creadas independientemente.
Filogenia molecular
La universalidad de la molécula portadora de la información genética hace que el DNA sea un carácter muy apropiado para el estudio comparativo y filogenético de las especies. Morfológicamente no es posible comparar una bacteria con un hombre, sin embargo si que es posible establecer una comparación con moléculas de DNA de ambos organismos, ya que están formadas por el mismo lenguaje de bases. Con datos de secuencias podemos comparar cualesquier grupo de organismos, por distantes que sean. Los datos moleculares tienen otras propiedades adicionales que todas juntas los convierten en el carácter ideal de estudios filogenéticos. Muchos trabajos obtienen y analizan las secuencias de genes y proteínas de diferentes especies para resolver cuestiones todavía dudosas de relaciones entre organismos. Los datos moleculares han demostrado que nuestra especie está mucho más cerca del chimpancé y el gorila de lo que creíamos.
Árbol filogenético Universal
El análisis molecular de secuencias también nos ha enseñado que hay una división en la raíz misma del árbol de la vida que es más fundamental que la división de 5 reinos que se enseña normalmente. En lugar de los dos tipos celulares canónicos, los procariotas y eucariotas, hay tres tipos principales de células, las arqueobacterias, las eubacterias y los eucariotas. Este nuevo árbol recibe el nombre de árbol filogenético universal.

Árbol filogenético universal
La sexta extinción en masa
El destino natural de cada especie es su extinción. Pero la vida continúa porque muchas especies dejan especies descendientes antes de morir. La diversidad biológica es un proceso dinámico que resulta del equilibrio entre la extinción y la producción de especies. Ninguna de los dos puede predominar por mucho tiempo. Una extinción que fuese superior a la producción de especies durante muchas generaciones conduciría a la pérdida de la vida sobre la Tierra, mientras que una situación inversa llevaría al agotamiento de los recursos y por tanto a la extinción. Actualmente desaparece una especie cada 15 minutos. El crecimiento desmesurado de la especie humana ha aumentado en mil veces la tasa normal de extinción, creando una situación que es análoga a la de una gran catástrofe. Estamos frente a la sexta, y quizá definitiva, gran extinción de la Tierra.
El movimiento creacionista
A pesar de la resistencia inicial a la visión darwinista, a principios del siglo XX la idea de la evolución de la vida era ya aceptada por la mayoría de las confesiones cristianas. Sin embargo, en el sur y el medio oeste de los EE.UU., numerosos grupos de cristianos evangélicos muy activos han intentado repetidamente prohibir la enseñanza de la evolución en las escuelas o compartirla en tiempo y reconocimiento científico con el relato bíblico del Génesis. En tiempos recientes, los creacionistas se han renovado y organizado a través del movimiento del diseño inteligente, ganando adeptos de otras confesiones. Los fundamentalistas dan un valor supremo a un texto escrito hace más de 2500 años en un contexto histórico y cultural que nada tiene que ver con el actual, y que aún así lo consideran más creíble que todo el cuerpo de conocimientos, datos empíricos y estructuras teóricas que millares de mentes dotadas de nuestra especie han desarrollado trabajando en colaboración durante los últimos 150 años. La universalización de la práctica de la ciencia no se ha traducido en la adopción de una cosmovisión común del Universo y la vida. La persistencia, un siglo y medio después del Origen de las especies, de creencias abiertamente anticientíficas es una gran paradoja de nuestro tiempo. La consecución de un credo universal de base darwiniana conduciría a un plano superior de conciencia de la especie humana y sería, sin duda, el mayor legado de Darwin. (Véase el artículo Darwinismo - Creacionismo escrito por el autor)
Los estudios y afirmaciones acerca de la evolución generalmente se refieren a uno de dos aspectos distintos: (1) las investigaciones acerca del hecho de la evolución y (2) las que se refieren al mecanismo de la evolución. Las primeras abarcan las disciplinas biológicas, tales como la paleontología, la clasificación, la biología comparada, la biología de poblaciones,... que muestran de manera inequívoca el hecho de la evolución. Las segundas, las afirmaciones acerca del mecanismo de la evolución, son el objeto principal de estudio de la genética de poblaciones, y nos informan de los factores, fuerzas o procesos que producen el cambio evolutivo, es decir, los mecanismos naturales que causan la descendencia con modificación. Una analogía cotidiana que ilustra esta distinción es la del tiempo meteorológico. Las precipitaciones, los vientos, las gotas frías, los tifones, son las evidencias que constituyen las afirmaciones de hecho del tiempo atmosférico. Ahora bien, si queremos explicar el porqué se dan los diferentes fenómenos meteorológicos, entonces nos tenemos que introducir en el ámbito de las afirmaciones del proceso o de los mecanismos meteorológicos. Debemos proponer los factores, tales como las diferencias de temperatura entre distintas masas de aire, que producen los fenómenos meteorológicos.
La evidencia de la evolución
La evolución que se da en una escala reducida, en el interior de una especie y en el intervalo de unas pocas generaciones, se denomina microevolución. La macroevolución es la evolución a gran escala, y abarca periodos considerables de tiempo, y grandes procesos de transformación; en el caso más extremo comprendería toda la evolución de la vida. Se pueden efectuar experimentos y/o observaciones de poblaciones de especies actuales a pequeña escala y obtener evidencia directa de evolución. Hay muchos ejemplos en los que se detecta la evolución en acción, como el clásico caso del melanismo industrial que se comentará más adelante. La selección artificial efectuada por el hombre en el perro o el caballo son también claros ejemplos que muestran el potencial de modificación de una especie. Por su propia dimensión temporal, no podemos demostrar la macroevolución directamente, exceptuando los casos de creación de nuevas especies de plantas mediante cruzamiento de especies distintas por el hombre. Aunque la evidencia evolutiva que suministra el registro fósil, la biología comparada, y la biología molecular es indirecta, no por ello es menos concluyente a la hora de demostrar la comunidad de origen de todos los organismos.
Las diferentes razas de perro que el hombre ha obtenido por selección artificial ilustran el potencial de cambio que tienen las especies.
El registro fósil
Los sedimentos que se han ido acumulando sobre la corteza de la tierra durante su historia geológica dejan una huella inestimable, generalmente en forma de huesos o esqueletos duros petrificados, de organismos muertos en el pasado: son los fósiles. El registro fósil es una ventana maravillosa a la historia de la vida. Si no existiera no podríamos inventarlo. Sin él, el vacío acerca de la evolución de la vida sobre la tierra sería insustituible. Podríamos especular, teorizar infinitamente, pero ¿quién podría haber imaginado que la Tierra estuvo dominada durante 150 millones de años por unos reptiles inmensos y fantásticos, los dinosaurios, que desaparecieron en un instante relativo de tiempo, si no hubieran existido fósiles de dinosaurios que nos lo contasen? La desintegración de los elementos químicos radiactivos que hay en las rocas ha permitido estimar que la Tierra se originó alrededor de hace 4600 millones de años. La Tierra, que era una esfera caliente, se enfría gradualmente, iniciándose un periodo de evolución química que culminará con la formación de las primeras células. En Australia y África se han encontrado sedimentos retenidos y fijados por bacterias de hace 3600 millones de años, lo que hace que esta fecha sea una estima mínima de la edad de inicio de la evolución biológica. La magnitud del tiempo en el que ha transcurrido la evolución se escapa completamente a nuestra comprensión, no podemos siquiera imaginar, limitados a la minúscula escala de nuestro tiempo vital, el potencial de transformación que suponen 3600 millones de años de evolución.
El tiempo geológico
El tiempo geológico se ha dividido en una serie de etapas jerárquicas, las eras, los períodos, y las épocas, que no siguen una cronología lineal, sino que es una crónica de los momentos claves de la historia de la vida. Las transiciones entre las cuatro eras, la Precámbrica, la Paleozoica, la Mesozoica y la Cenozoica representan grandes cambios en las fauna y flora de toda la Tierra. En el primer período de la era Paleozoica, el Cámbrico, hace 570 millones de años, aparecen de golpe en el registro fósil los animales pluricelulares que tienen partes duras, como las conchas, y exoesqueletos,... El final del Paleozoico coincide con la mayor extinción habida en la Tierra, en la que desaparecieron el 96% de las especies. Al final del Mesozoico, en la transición entre el período Cretácico y Terciario, se da la conocida extinción de los dinosaurios, junto a un 70% de las especies existentes.
Cronología de siete momentos estelares de la evolución
¿Qué nos enseña el registro fósil sobre la historia de la vida sobre la Tierra? Esta es una lista de los acontecimientos más importantes
- Origen de la célula procariota 3600 M (M=Millones de años)
- Origen de la célula eucariota 1400 M
- Origen de la fauna de animales pluricelulares 650 M
- Fauna de la explosión cámbrica 570 M
- Origen de los vertebrados terrestres 360 M
- Extinción de los dinosaurios. La antorcha pasa de los dinosaurios a los mamíferos 65 M
- Origen de Homo sapiens 0,1 M.
Si toda la historia de la Tierra la comprimiésemos en una hora, a los 20 minutos aparecerían las bacterias, a los 55 los dinosaurios, los antropoides aparecen a 40 segundos antes del final, y los humanos al cumplirse la hora.
Hay una coherencia entre el registro fósil y el origen evolutivo de las especies y sus grupos correspondientes. Si la vida se debiera a la creación espontánea e independiente de especies, no tendríamos porqué esperar que el hombre surgiera con los mamíferos, podríamos haber surgido en cualquier momento, cuando aparecieron los peces, o los dinosaurios, o antes de que surgieran los primates.
La evolución no tiene dirección definida
La historia de la vida es una historia de extinciones y muerte,... con unos pocos supervivientes. El 99,9% de las especies que han existido alguna vez están hoy extintas. Grupos enteros de organismos, como los dinosaurios (exceptúando las aves), los trilobites, los ammonoideos, se han extinguido para siempre sin dejar descendiente alguno.
Como señala el reconocido paleontólogo S. Gould, el registro fósil no es un relato convencional que conduce a los diferentes linajes a más excelencia, más complejidad, más diversidad. La historia de la vida no muestra un rumbo definido, no tiene dirección ni sentido. La evolución es una narración de eliminación masiva seguida de diferenciación en el interior de unos cuantos supervivientes. Es a priori imposible determinar la dirección de la evolución porque la importancia de los acontecimientos concretos, contingentes, como la extinción o no de un grupo de organismos en el caso de una extinción en masa, o la posesión o no de una variante adaptativa adecuada cuando ésta es requerida, son los verdaderos agentes de la historia.
Las extinciones en masa
Hay dos regímenes de extinciones: la extinción normal, que afecta a las especies que dejan de seguir a su ambiente en su lucha diaria por adaptarse, y la extinción en masa, que son más rápidas y devastadoras en cuanto a su magnitud. Al menos han ocurrido cinco extinciones en masa, y han dejado muchos huecos ecológicos que han permitido que sean ocupados por los descendientes de las especies supervivientes. Esta ocupación del bioespacio disponible suele ir acompañada de una rápida y extensa diversificación morfológica que se denomina radiación adaptativa. No se sabe con certeza la causa de las extinciones masivas, aunque las causas físicas como el impacto de asteroides o el cambio climático parecen más probables que las biológicas. Según una hipótesis reciente, hay ciclos de extinción masiva aproximadamente cada 26 millones de años, y el impacto periódico de lluvias de cometas sobre la Tierra podría explicar dicho ciclo.
La fauna de Burguess Shale
Si tuviéramos la oportunidad de volver a un momento del pasado, el período previo al Cámbrico sería sin duda uno de los más atractivos. En el Cámbrico, hace 570 M, se produce la gran explosión en el registro fósil de los primeros animales pluricelulares con partes duras. Darwin se preguntaba por qué estos primeros animales eran ya anatómicamente complejos y sin precursores aparentes. La respuesta está en el yacimiento de Burguess Shale, situado en las montañas rocosas canadienses. Debido a condiciones muy especiales de conservación, aquí se encuentra la única fauna de cuerpo blando (sin estructuras duras) que existe de un tiempo inmediatamente anterior a la explosión cámbrica.
La gran sorpresa de Burguess Shale es que las pocas especies analizadas contienen una disparidad de diseños anatómicos que exceden, con mucho, la gama moderna que hay en todo el mundo. En ninguna fauna posterior se repite la riqueza anatómica de este yacimiento. De los 120 géneros analizados, 20 son diseños de artrópodos únicos, y además de estar representados los 4 grupos de animales que hoy existen, hay 8 diseños que no encajan en ningún grupo animal conocido. La época de Burguess Shale parece ser que fue un tiempo asombroso de experimentación, una era de gran flexibilidad evolutiva, que fue seguido de una gran extinción.
Opabinia: una vida extraña de un remoto pasado
Opabinia fue el fósil de Burguess Shale que reveló la caja de sorpresas que había en este yacimiento. Su diseño singular no pertenece a ningún grupo animal conocido. ¡Tiene 5 ojos!, trompa flexible formada por un tubo cilíndrico estriado, un tubo digestivo en forma de U, un tronco de 15 segmentos, con lóbulos delgados laterales. Los 3 segmentos últimos forman una cola con tres pares de hojas delgadas y lobuladas dirigidas hacia afuera.
Opabinia fue el primer fósil del yacimiento de Burguess Shale que demostró la singularidad morfológica de las primeras formas pluricelulares. Tiene 5 ojos, trompa flexible, tubo digestivo en forma de U, un tronco de 15 segmentos, los 3 últimos forman una cola.
Los dinosaurios
Durante el periodo Triásico surgen los dinosaurios de una línea de los reptiles e inician una dinastía sobre el medio terrestre que habría de durar 150 M. Se han identificado más de 350 especies de dinosaurios, y se cree que esta es una estima muy inferior a la de las especies que realmente existieron. Entre los dinosaurios están los animales más grandes que alguna vez hayan vivido sobre la Tierra. El Seismosaurus, el mayor herbívoro conocido, tenía unas 40 metros de longitud. El Tyrannosaurus rex, uno de los mayores carnívoros, tenía 12 m de longitud. Los dinosaurios se dividen en dos grandes órdenes, los de pelvis de lagarto (Saurisquios), que comprenden tanto especies carnívoras como herbívoras, y los de pelvis de ave (Ornistiquios), cuyas especies eran todas herbívoras. Los dinosaurios se extinguieron casi en su totalidad, exceptuando una línea menor que dio lugar a las aves, hace 65 M. Junto a ellos, desaparecieron en un breve lapso de tiempo el 75% de las especies existentes. Se ha propuesto que el impacto de un asteroide sobre la superficie terrestre podría ser la causa de la extinción en masa.
¿Quién podría haber imaginado que la Tierra estuvo dominada durante 150 millones de años por unos reptiles inmensos y fantásticos, los dinosaurios, que desaparecieron en un instante relativo de tiempo, si no hubieran existido fósiles de dinosaurios que nos lo contasen?
Biología comparada: homología y analogía
Cuando uno observa similitudes entre especies, se pueden distinguir entre dos tipos de semejanzas, la analogía y la homología. El ala de un ave y el de una mosca forman una extensión plana y tienen un movimiento de aleteo similar; los peces, los delfines, o los pingüinos tienen una sección transversal aplanada que les permite desplazarse por el agua. Estas semejanzas, llamadas analogías, son más bien superficiales y se deben a que estos organismos están sometidos a las mismas restricciones funcionales o adaptativas, y no son debidas a que posean un antepasado común reciente.
En contraste con la analogía, una homología es la similitud que hay entre caracteres de distintas especies debido a que tienen un origen común, y no a la acción directa de una presión funcional. Por ejemplo, todos los tetrápodos (animales vertebrados terrestres con cuatro extremidades) tienen la extremidad de cinco dedos, y ésta se encuentra tanto en las alas de la aves y de murciélagos como en la mano del ser humano, a pesar que estas extremidades representan unos papeles funcionales muy distintos. La razón de esta estructura común es que todos los tetrápodos conservamos la misma estructura básica de la especie ancestral original.
Homología: todos los tetrápodos tienen una extremidad con cinco dedos, aunque tengan diferentes funciones.
La homología es la base de la clasificación
La clasificación se basa en la comparación de los caracteres de las especies, y los caracteres homólogos son los elementos claves para establecer una clasificación evolutiva. Si las especies proceden de otras especies por evolución, y además no varían tan rápidamente como para perder toda su herencia histórica, se esperaría que los distintos seres vivos compartieran una serie de caracteres homólogos. El análisis de los diferentes caracteres fenotípicos, como la morfología, la conducta, los cromosomas, la anatomía externa e interna, el desarrollo embrionario, el metabolismo, la variación genética y proteica muestran que las especies presentan semejanzas homológas en todos los niveles del fenotipo. Cuanto más próxima sean la especies, mayor será el grado de semejanza, y lo contrario también es cierto, cuanto más alejada estén menos semejanzas encontraremos. Así, las diferencias que hoy vemos entre las especies se deben a las nuevas variaciones que han adquirido desde su separación del antepasado común. Las similitudes que atribuimos a las homologías no podrían explicarse si las especies se originasen independientemente unas de otras.
Órganos vestigiales
Un caso especialmente significativo de homología es la de los órganos vestigiales o residuales. La pelvis de los tetrápodos es una estructura ósea cuya función es articular las extremidades posteriores. Las ballenas son tetrápodos cuyos antepasados mamíferos abandonaron la tierra para habitar en el mar. En este nuevo medio las ballenas perdieron sus extremidades inferiores, pero aún conservan como huella acusatoria de su pasado tetrápodo, la serie completa de los huesos de la pelvis. De forma similar, las serpientes presentan vestigios de la extremidad posterior que portaban sus antepasados.
La reducida pelvis de los cetáceos es un órgano vestigial que prueba su origen tetrápodo.
Biología molecular y homología
La biología molecular ha suministrado la evidencia más universal de homología. Todos los organismos vivos compartimos el mismo material hereditario, el DNA, una molécula helicoidal cuya información se encuentra codificada en 4 letras o nucleótidos distintos. Igualmente, el código genético es prácticamente Universal, todos los organismos comparten el mismo diccionario que da el significado a la secuencia de DNA. Ambos ejemplos son pruebas muy robustas de la relación íntima que existe entre lo viviente.
El DNA es una molécula helicoidal que tiene información genética codificada a partir de cuatro letras o nucleótidos distintos.
El árbol de la vida: filogenia
Si la historia de la vida es cambio y ramificación por descendencia, entonces su representación sería la de un árbol o filogenia, en la que el tronco y las ramas internas se corresponderían a los antepasados de las especies actuales y los extremos de las ramas externas serían las especies actuales. ¿Como se establece una filogenia? Ordenando las especies actuales atendiendo a la similaridad morfológica y/o genética de sus caracteres homólogos.
El sistema linneano
El botánico sueco Carolus Linnaeus (1707-78) ideó el sistema de clasificación que se utiliza, con algunas modificaciones, hoy en día. Hay siete niveles inclusivos de clasificación, que son, de menor a mayor, la especie, el género, la familia, el orden, la clase, el tipo o Phylum y el reino. El nombre científico de cada especie tiene dos partes, el león, por ejemplo, se denomina Panthera leo. La primera parte se refiere al género y la segunda a la especie. Consideremos un ejemplo de como se agrupan las especies actuales en las distintas categorías linneanas. El león, la pantera, el tigre, pertenecen al género Panthera, que junto al género Felix (el gato doméstico) y otros se agrupan en la familia de los felinos. Los felinos, con los cánidos y úrsidos, constituyen el orden de los carnívoros. Primates, roedores, carnívoros, ... se reúnen en la clase mamíferos. Estos organismos comparten características como el amamantar a sus crías con leche, gestarlas en el útero mediante un órgano complejo, la placenta. Su piel está protegida por pelaje o pelos. Mamíferos, aves, reptiles, anfibios, y peces se reúnen en un solo tipo o phylum, porque todos tienen columna vertebral, un máximo de cuatro miembros, y sangre roja con hemoglobina, son los cordados. Los insectos, las arañas, los crustáceos, y los ciempiés, se clasifican en otro tipo, los artrópodos. Las almejas, caracoles y calamares se agrupan en los moluscos, y así sucesivamente. El tipo cordado, artrópodo, molusco y otros forman el reino Animal. El árbol de la vida se ordena siguiendo divisiones que van de características generales a aspectos más específicos.
El botánico sueco Carolus Linnaeus (1707-78) creó el sistema de clasificación que se utiliza hoy en día.
Significado de los niveles jerárquicos de clasificación
¿Por qué se ordena la diversidad biológica en diferentes niveles jerárquicos? Los rasgos de las divisiones más generales corresponderían a adaptaciones básicas o principales que surgieron en los momentos iniciales de la evolución de los especies progenitoras de estos grupos. Por ejemplo, hay cinco grandes reinos, las moneras, los protistas, los hongos, las plantas y los animales, que se corresponden con las cinco diferenciaciones principales de la vida sobre la Tierra. El Phylum o tipo es la siguiente unidad básica de diferenciación entre reinos, y podrían entenderse como proyectos básicos fundamentales de anatomía. En el reino Animal estos planos fundamentales serían las esponjas, los anélidos (gusanos), los artrópodos (insectos, crustáceos,...), los cordados, .... A partir de estas adaptaciones principales surgen posteriormente nuevas variaciones o subadaptaciones diversas, que por tener un menor tiempo de evolución, tienen un rango clasificatorio menor.
La clasificación de la diversidad biológica es una prueba de la evolución
El hecho que la diversidad de la vida esté jerarquizada es un fuerte argumento en favor de la evolución. El que las especies compartan estructuras anatómicas y adaptaciones básicas puede explicarse fácilmente si suponemos que las especies actuales compartieron antepasados, pero no esperaríamos dicho patrón si las especies hubieran sido creadas independientemente.
Filogenia molecular
La universalidad de la molécula portadora de la información genética hace que el DNA sea un carácter muy apropiado para el estudio comparativo y filogenético de las especies. Morfológicamente no es posible comparar una bacteria con un hombre, sin embargo si que es posible establecer una comparación con moléculas de DNA de ambos organismos, ya que están formadas por el mismo lenguaje de bases. Con datos de secuencias podemos comparar cualesquier grupo de organismos, por distantes que sean. Los datos moleculares tienen otras propiedades adicionales que todas juntas los convierten en el carácter ideal de estudios filogenéticos. Muchos trabajos obtienen y analizan las secuencias de genes y proteínas de diferentes especies para resolver cuestiones todavía dudosas de relaciones entre organismos. Los datos moleculares han demostrado que nuestra especie está mucho más cerca del chimpancé y el gorila de lo que creíamos.
Árbol filogenético Universal
El análisis molecular de secuencias también nos ha enseñado que hay una división en la raíz misma del árbol de la vida que es más fundamental que la división de 5 reinos que se enseña normalmente. En lugar de los dos tipos celulares canónicos, los procariotas y eucariotas, hay tres tipos principales de células, las arqueobacterias, las eubacterias y los eucariotas. Este nuevo árbol recibe el nombre de árbol filogenético universal.
Árbol filogenético universal
La sexta extinción en masa
El destino natural de cada especie es su extinción. Pero la vida continúa porque muchas especies dejan especies descendientes antes de morir. La diversidad biológica es un proceso dinámico que resulta del equilibrio entre la extinción y la producción de especies. Ninguna de los dos puede predominar por mucho tiempo. Una extinción que fuese superior a la producción de especies durante muchas generaciones conduciría a la pérdida de la vida sobre la Tierra, mientras que una situación inversa llevaría al agotamiento de los recursos y por tanto a la extinción. Actualmente desaparece una especie cada 15 minutos. El crecimiento desmesurado de la especie humana ha aumentado en mil veces la tasa normal de extinción, creando una situación que es análoga a la de una gran catástrofe. Estamos frente a la sexta, y quizá definitiva, gran extinción de la Tierra.
El movimiento creacionista
A pesar de la resistencia inicial a la visión darwinista, a principios del siglo XX la idea de la evolución de la vida era ya aceptada por la mayoría de las confesiones cristianas. Sin embargo, en el sur y el medio oeste de los EE.UU., numerosos grupos de cristianos evangélicos muy activos han intentado repetidamente prohibir la enseñanza de la evolución en las escuelas o compartirla en tiempo y reconocimiento científico con el relato bíblico del Génesis. En tiempos recientes, los creacionistas se han renovado y organizado a través del movimiento del diseño inteligente, ganando adeptos de otras confesiones. Los fundamentalistas dan un valor supremo a un texto escrito hace más de 2500 años en un contexto histórico y cultural que nada tiene que ver con el actual, y que aún así lo consideran más creíble que todo el cuerpo de conocimientos, datos empíricos y estructuras teóricas que millares de mentes dotadas de nuestra especie han desarrollado trabajando en colaboración durante los últimos 150 años. La universalización de la práctica de la ciencia no se ha traducido en la adopción de una cosmovisión común del Universo y la vida. La persistencia, un siglo y medio después del Origen de las especies, de creencias abiertamente anticientíficas es una gran paradoja de nuestro tiempo. La consecución de un credo universal de base darwiniana conduciría a un plano superior de conciencia de la especie humana y sería, sin duda, el mayor legado de Darwin. (Véase el artículo Darwinismo - Creacionismo escrito por el autor)
Evolución humana
Las semejanzas morfológicas, bioquímicas, y genéticas sitúan al ser humano en el orden de los primates de la clase mamíferos. Dentro de los primates, son el chimpancé, el gorila y el orangután, sus parientes más próximos. Los datos de comparación de secuencias muestran que hay una similitud del 98,5% entre el DNA humano y el del chimpancé. Esta semejanza es mayor que la que existe entre el chimpancé y el gorila o el gorila y nosotros, por lo que el chimpancé y los humanos compartimos un antecesor común más reciente que ambos con los gorilas. Esta cercanía, que se ha estimado en 5M de años, es mucho mayor de lo que se había inferido sólo con datos morfológicos, y muestra la capacidad de los datos de DNA para desvelar relaciones de parentesco. En la evolución humana existen dos grandes adquisiciones, la marcha bípeda, y el desarrollo extraordinario del cerebro. El registro fósil nos muestra que la postura erguida precedió al desarrollo cerebral y que África es la cuna de la humanidad. El Australopithecus, de una antigüedad de 1,5-5M de años es el primer mono antropoide de marcha bípeda. Su capacidad craneal era similar a la del chimpancé y gorila actual. El Homo habilis y el Homo erectus son las líneas que siguen cronológicamente hasta la llegada de nuestra especie, Homo sapiens, hace 100.000 años.


Filogenia actual del humanos y antropomorfos modernos que integra los datos moleculares y morfológicos. H: hombre, C: Chimpancé, G: Gorila, O: Orangután y G: Gibón.
DNA del hombre de Neandertal
En 1997 se publica la secuencia de un trozo del DNA del primer fósil que se encontró del hombre de Neandertal, una subespecie extinta de la especie humana. Es la primera vez que se obtiene la secuencia molecular de un fósil humano. Cuando la secuencia se comparó con secuencias homólogas de DNA humano actual se dedujo que el antepasado común de nosotros y el hombre de Neandertal vivió hace 500.000 años, de lo que se concluyó que el hombre de Neandertal se extinguió sin mezclarse con el hombre actual.
P.S.: La publicación de la secuencia completa del genoma de Neandertal en mayo de 2010 ha revelado que hubo cierto flujo genético tras un primer contacto inicial en Asia Menor, de modo que entre el 1-4% del genoma de la humanidad actual (exceptuando los africanos subsaharianos) procede del hombre de Neandertal.
 El análisis de la secuencia genómica de DNA del hombre de Neandertal indica que entre el 1-4% del genoma de la humanidad actual (exceptuando los africanos subsaharianos) procede de esta especie humana.
El análisis de la secuencia genómica de DNA del hombre de Neandertal indica que entre el 1-4% del genoma de la humanidad actual (exceptuando los africanos subsaharianos) procede de esta especie humana.
El futuro de la evolución

Con la aparición del cerebro humano se produce un hecho singular: el surgimiento de la conciencia objetiva en la biosfera. Somos chispas de conciencia en un Universo que hasta nuestra irrupción era ciego y sordo a su devenir. Por primera vez, y en un sentido recursivo, el universo se piensa a si mismo cuando descubrimos su existencia de igual forma que nosotros nos pensamos y descubrimos con nuestro cerebro. Esto constituye el momento más trascendental de toda la evolución. El pensamiento científico que resulta de la actividad de mentes que trabajan en colaboración para entender la realidad física, biológica y mental de nuestro universo nos conduce a niveles de conciencia superiores. Como señala Mosterín (2005), nos dirigimos hacia a una conciencia cósmica, donde se alcanzarán planos superiores de empatía, alegría y lucidez.
La evolución científica y cultural no se limita a las leyes de la selección natural y la herencia genética. Sigue un proceso de transmisión horizontal (entre individuos de una generación) y vertical (entre generaciones) que es mucho más veloz que los procesos de evolución biológica típicos. El ser humano está en el umbral de poder dirigir la evolución en la dirección que el crea conveniente. Podrá limitar enormemente los azares de la mutación y de la segregación genética y unión de gametos. Genes mutados que causan graves enfermedades a la especie humana podrán ser sustituidos por sus contrapartidas no deletéreas,... Pero paradójicamente la humanidad se enfrenta hoy a los retos de su propio éxito evolutivo. El crecimiento explosivo de su población, con las necesidades de espacio y recursos que genera, provoca la eliminación o reducción hasta tamaños insoportables de los hábitats de las especies. La tasa de extinción actual no es sostenible. Es posible que el ser humano pueda mantener la diversidad biológica en forma de semillas o células congeladas, e incluso que pueda compensar la pérdida de especies con la creación de nuevas mediante ingeniería genética. Pero la reducción drástica de espacios naturales limitará inevitablemente la diversidad. Los grandes retos de la humanidad son el control del crecimiento de la población, la eliminación de las desigualdades socioeconómicas, el mantenimiento de un desarrollo sostenido y viable y la conservación de hábitats naturales y de especies. El éxito frente a tales desafíos precisa del desarrollo de una conciencia ética universal basada en el respeto a la diversidad de pueblos y culturas de nuestra especie. Dicha conciencia debe extenderse hasta abarcar a la totalidad de la vida, al conjunto precioso de formas orgánicas que nos han acompañado, desde la primera célula antecesora común, en este fascinante periplo singular e irrepetible que es nuestra historia biológica.
Semana del 31 de enero al 3 de febrero del 1017
El código genético
ve al siguiente link, observa el vídeo y sigue las flechas para que al final puedas resolver la pregunta en el cuaderno. La cual se sustentara en la clase
Link: Contenido material genetico
o copia y pega el siguiente link en la barra de dirección
http://contenidosparaaprender.mineducacion.gov.co/G_8/S/S_G08_U02_L07/S/S_G08_U02_L07/S_G08_U02_L07_03_06.html
 ----------------- 2016 -----------------
----------------- 2016 -----------------
Las semejanzas morfológicas, bioquímicas, y genéticas sitúan al ser humano en el orden de los primates de la clase mamíferos. Dentro de los primates, son el chimpancé, el gorila y el orangután, sus parientes más próximos. Los datos de comparación de secuencias muestran que hay una similitud del 98,5% entre el DNA humano y el del chimpancé. Esta semejanza es mayor que la que existe entre el chimpancé y el gorila o el gorila y nosotros, por lo que el chimpancé y los humanos compartimos un antecesor común más reciente que ambos con los gorilas. Esta cercanía, que se ha estimado en 5M de años, es mucho mayor de lo que se había inferido sólo con datos morfológicos, y muestra la capacidad de los datos de DNA para desvelar relaciones de parentesco. En la evolución humana existen dos grandes adquisiciones, la marcha bípeda, y el desarrollo extraordinario del cerebro. El registro fósil nos muestra que la postura erguida precedió al desarrollo cerebral y que África es la cuna de la humanidad. El Australopithecus, de una antigüedad de 1,5-5M de años es el primer mono antropoide de marcha bípeda. Su capacidad craneal era similar a la del chimpancé y gorila actual. El Homo habilis y el Homo erectus son las líneas que siguen cronológicamente hasta la llegada de nuestra especie, Homo sapiens, hace 100.000 años.
Filogenia actual del humanos y antropomorfos modernos que integra los datos moleculares y morfológicos. H: hombre, C: Chimpancé, G: Gorila, O: Orangután y G: Gibón.
DNA del hombre de Neandertal
En 1997 se publica la secuencia de un trozo del DNA del primer fósil que se encontró del hombre de Neandertal, una subespecie extinta de la especie humana. Es la primera vez que se obtiene la secuencia molecular de un fósil humano. Cuando la secuencia se comparó con secuencias homólogas de DNA humano actual se dedujo que el antepasado común de nosotros y el hombre de Neandertal vivió hace 500.000 años, de lo que se concluyó que el hombre de Neandertal se extinguió sin mezclarse con el hombre actual.
P.S.: La publicación de la secuencia completa del genoma de Neandertal en mayo de 2010 ha revelado que hubo cierto flujo genético tras un primer contacto inicial en Asia Menor, de modo que entre el 1-4% del genoma de la humanidad actual (exceptuando los africanos subsaharianos) procede del hombre de Neandertal.
El análisis de la secuencia genómica de DNA del hombre de Neandertal indica que entre el 1-4% del genoma de la humanidad actual (exceptuando los africanos subsaharianos) procede de esta especie humana.
El futuro de la evolución
Con la aparición del cerebro humano se produce un hecho singular: el surgimiento de la conciencia objetiva en la biosfera. Somos chispas de conciencia en un Universo que hasta nuestra irrupción era ciego y sordo a su devenir. Por primera vez, y en un sentido recursivo, el universo se piensa a si mismo cuando descubrimos su existencia de igual forma que nosotros nos pensamos y descubrimos con nuestro cerebro. Esto constituye el momento más trascendental de toda la evolución. El pensamiento científico que resulta de la actividad de mentes que trabajan en colaboración para entender la realidad física, biológica y mental de nuestro universo nos conduce a niveles de conciencia superiores. Como señala Mosterín (2005), nos dirigimos hacia a una conciencia cósmica, donde se alcanzarán planos superiores de empatía, alegría y lucidez.La evolución científica y cultural no se limita a las leyes de la selección natural y la herencia genética. Sigue un proceso de transmisión horizontal (entre individuos de una generación) y vertical (entre generaciones) que es mucho más veloz que los procesos de evolución biológica típicos. El ser humano está en el umbral de poder dirigir la evolución en la dirección que el crea conveniente. Podrá limitar enormemente los azares de la mutación y de la segregación genética y unión de gametos. Genes mutados que causan graves enfermedades a la especie humana podrán ser sustituidos por sus contrapartidas no deletéreas,... Pero paradójicamente la humanidad se enfrenta hoy a los retos de su propio éxito evolutivo. El crecimiento explosivo de su población, con las necesidades de espacio y recursos que genera, provoca la eliminación o reducción hasta tamaños insoportables de los hábitats de las especies. La tasa de extinción actual no es sostenible. Es posible que el ser humano pueda mantener la diversidad biológica en forma de semillas o células congeladas, e incluso que pueda compensar la pérdida de especies con la creación de nuevas mediante ingeniería genética. Pero la reducción drástica de espacios naturales limitará inevitablemente la diversidad. Los grandes retos de la humanidad son el control del crecimiento de la población, la eliminación de las desigualdades socioeconómicas, el mantenimiento de un desarrollo sostenido y viable y la conservación de hábitats naturales y de especies. El éxito frente a tales desafíos precisa del desarrollo de una conciencia ética universal basada en el respeto a la diversidad de pueblos y culturas de nuestra especie. Dicha conciencia debe extenderse hasta abarcar a la totalidad de la vida, al conjunto precioso de formas orgánicas que nos han acompañado, desde la primera célula antecesora común, en este fascinante periplo singular e irrepetible que es nuestra historia biológica.
Semana del 31 de enero al 3 de febrero del 1017
ve al siguiente link, observa el vídeo y sigue las flechas para que al final puedas resolver la pregunta en el cuaderno. La cual se sustentara en la clase
Link: Contenido material genetico
o copia y pega el siguiente link en la barra de dirección
http://contenidosparaaprender.mineducacion.gov.co/G_8/S/S_G08_U02_L07/S/S_G08_U02_L07/S_G08_U02_L07_03_06.html

¡BIENVENIDOS!
ESTE ES EL MARAVILLOSO MUNDO DE LA
BIOLOGÍA.
ESTE ES UN ESPACIO ESPECIALMENTE DISEÑADO PARA USTEDES. AQUÍ PODRÁN ENCONTRAR INFORMACIÓN MUY IMPORTANTE PARA ENRIQUECER SU CONOCIMIENTO. CON EL FIN DE APOYAR SUS PROCESOS ACADÉMICOS SE SUBIRÁ MATERIAL DIDÁCTICO Y, ADEMAS, SE DEJARAN ACTIVIDADES Y MUCHÍSIMOS RECURSOS MAS.

Maria Tereza Carmona duque
biology teacher
mariacarmona@windsorroyalschool.edu.co
_________________________________________________________________________________
_________________________________________________________________________________
TALLER PARA ENTREGAR EL DÍA 17 DE AGOSTO
EN EL CUADERNO
REPRODUCCIÓN HUMANA
Seré exigente con los dibujos y diagramas
- ¿Que es la menstruación y que se expulsa en ella?
- ¿En que parte del aparato reproductor femenino ocurre la fecundación?
- ¿Cuales son las gónadas femeninas y cuales las masculinas?
- ¿Cuales son los gametos femeninos y masculinos?
- ¿Cuales son las partes del espermatozoide Dibújalas y explica la función de cada una
- ¿Que nombre recibe la capa del útero donde se implantara el embrión ?
- ¿Que función sexual reproductiva cumplen las feromonas?
- ¿Cual es la hormona que regula los caracteres sexuales masculinos?
- ¿Cual es la hormona que regula los caracteres sexuales femeninos?
- ¿Que función cumplen los estrogenos y como actúan en la anticoncepción?
- ¿Como y cuando ocurre la ovulación y?
- Dibuja los aparatos reproductores femeninos y masculinos con sus respectivas partes.
TEMAS DEL 1 AL 12 DE AGOSTO
La reproducción sexual en animales
Todos los organismos animales proceden del desarrollo de una célula llamada huevo o cigoto que procede de la unión de los gametos, un espermatozoide y un óvulo, células especiales que se forman en las gónadas, testículos y ovarios respectivamente.
En los animales existe una gran diversidad de formas de reproducción sexual, la mayoría son unisexuales pero algunos grupos son hermafroditas como determinados anélidos, moluscos o peces, realizando una fecundación cruzada entre dos organismos, ya que no se pueden fecundar a sí mismos.
ENLACE IMPORTANTE
La reproducción asexual en los animales
|
| Animales sésiles que se reproducen asexualmente |
Tipos de reproducción asexual
La reproducción asexual ocurre sin la intervención de órganos sexuales especiales, produciéndose nuevos individuos a partir de un solo progenitor. En las plantas se presenta mediante la producción de esporas o bulbos. En los animales protistas, en gusanos poliquetos y planarias, y en animales sésiles acuáticos como los tunicados, cnidarios y esponjas la reproducción ocurre de distintas maneras.
Este tipo de reproducción puede llevarse a cabo por:
- Escisión unicelular - un animal se divide en mitades generalmente iguales que se desarrollan hasta formar dos adultos idénticos al original. En este proceso de división primero se divide el núcleo y luego el citoplasma. Los esporozoos, como el Paramecio, llevan a cabo este tipo de reproducción.
|
| Paramecio en proceso de división |
- Esporulación - en este caso el núcleo se divide varias veces y el citoplasma también. Cada pedazo de núcleo se lleva una porción del citoplasma. De cada división surge un nuevo individuo.
- Gemación - en este tipo de reproducción los nuevos individuos nacen en forma de yema, adheridos al original, formando una colonia. Ejemplos de animales que se reproducen así son: los tunicados y cnidarios como la hidra.
|
| Por gemación la hidra ha creado un nuevo individuo. |
- Fragmentación - esta forma de reproducción es algo curiosa. El individuo original se divide en varias partes y de cada parte surge un individuo nuevo idéntico al original. Algunos animales en los que se da este proceso son los nemertinos (gusanos) y turbelarios (planarias o gusanos planos).
|
| Nacimiento de una nueva estrella de mar por el proceso de fragmentación |
|
| De este gusano partido en dos partes surgirán dos gusanos iguales al original, regenerando la parte faltante. |
Ventajas de la reproducción asexual
- No necesita de una pareja para llevarse a cabo la reproducción.
- No requieren de una madurez sexual que los animales más evolucionados si necesitan.
- La reproducción puede ocurrir en cualquier momento.
- Es un proceso simple y sencillo.
- Se puede multiplicar la población rápidamente.
- Un individuo tiene mayor oportunidad de supervivencia de la especie de quedar aislado.
Desventajas de la reproducción asexual
- No hay un aporte genético de dos padres, por lo tanto el resultado es... copias iguales al original. Esto implica que no hay un aporte de información genética nueva para la evolución de los nuevos individuos.
Sea sexual o asexual la reproducción de los animales, es un proceso de importancia vital para su supervivencia
SEMANA DEL 25 AL 29 DE JULIO
ESCRIBE LAS SIGUIENTES PREGUNTAS!
1) ¿ Cómo es la reproducción asexual o vegetativa de las plantas?
2) ¿ Cuándo se da la reproducción sexual en las plantas?
3) ¿ Cómo ocurre la polinización.?
4) ¿ Cúales son los animales encargados de la polinización?
5) ¿ Qué son los estambres y los pistilos en las flores?
6) ¿ Por que se le da el nombre de hermafroditas a algunas flores?
7) Realiza los dibujos correspondientes
8) ¿Que significa que una planta es dioica o monoica?
RESULTADOS
ESTUDIANTES QUE NO DEBEN PRESENTAR ACTIVIDADES DE NIVELACIÓN PARA EL SEGUNDO PERIODO
MIGUEL FUENTES
JOSE PADILLA
JUAN DIEGO
JUAN CAMILO SALAZAR
DANIELA VILLALBA
LOS DEMÁS DEBEN PRESENTAR LA SIGUIENTE ACTIVIDAD DE NIVELACIÓN: EVALUACIÓN DE LOS TEMAS VISTOS EN EL SEGUNDO PERIODO
DÍA DEL EXAMEN JULIO 12 DEL 2016
Reproducción asexual de las plantas
Existen dos tipos de reproducción asexual en plantas: multiplicación vegetativa y por gérmenes:
1. Multiplicación vegetativa: Asegura la perpetuación de individuos bien adaptados a ese medio y evolutivamente eficaces. Es muy común incluso en plantas superiores. Existen dos tipos: la fragmentación y la división celular que engloba la bipartición y la gemación.
La fragmentación consiste la fragmentación de partes de células, talos o vástagos de los que surgen individuos hijos. Un ejemplo de fragmentación son los esquejes. En la bipartición, la célula madre se divide por completo en dos células hijas nuevas de igual tamaño. En la gemacióncelular el tamaño de la célula hija es al principio menor que el de la célula madre.
2. Por gérmenes. Los gérmenes son células asexuales reproductivas que desarrollan directamente el individuo. Existen varios tipos: pluricelulares -los propágulos- y generalmente unicelulares -las esporas-.
Hay zonas en que porciones del talo o del tallo de las plantas pluricelulares están particularmente especializadas para separarse de la planta madre y extenderse, son los propágulos (agrupaciones de células), son muy comunes en las plantas inferiores. Existen varios tipos, los hormogonios de las cianobacterias, los tubérculos de la patata, los dientes del ajo…
Las esporas son células germinales especialmente diferenciadas para la reproducción asexual.
Esporas
Son la forma más corriente de reproducción asexual en plantas, producen en general poca variabilidad, son agentes de dispersión y normalmente unicelulareas aunque hay esporas con varias células o núcleos.
Existen varios tipos de esporas según las condiciones de formación:
- Según la situación: exósporas o conidios si se forman al exterior por estrangulación y endósporas si se forman en el interior de un esporangio.
- Según la capacidad de dispersión: aplanósporas si son inmóviles como el polen, muchosconidios y zoosporas o planósporas si son móviles.
- Según la formación: mitósporas o neutrósporas si son diploide y meiosporas, gonosporas o esporas “sexuales” si son haploides.
Las esporas tienen también nombres especiales como por ejemplo: diplosporas si son diploide, haplósporas si son haploides, si son esporas de resistencia se las llama clamidosporas. Si se producen en ascas son ascosporas, basidiosporas si se producen en basidios. Heterósporas si son distintas generalmente de tamaño, micrósporas si son pequeñas y masculinas, megásporas si son grandes y femeninas.
Las estructuras especializadas donde se producen las esporas son los esporangios. Son unicelulares (sin cubierta) en algas y hongos; pluricelulares (con cubierta y arquesporio que es el tejido fértil) de briófitos a espermatófitos. Su nomenclatura es igual que la de las esporas, por ejemplo de meióspora meiosporangio.
Reproducción sexual
Este tipo de reproducción es un mecanismo mucho más complicado que la reproducción asexual y en ella intervienen muchos factores.
La reproducción sexual implica la unión de células germinales especiales, los gametos, y está encaminada a la variabilidad genética por recombinación cromosómica. Este proceso se realiza en varias etapas. Primero se realiza la meiosis para transformar las células diploides en haploides que son los gametos.
Posteriormente se produce la singamia o unión de gametos haploides para formar un zigoto diploide, que implica una plasmogamia (unión de citoplasmas) y una cariogamia o fecundación (unión de núcleos).
Los gametos suelen ser haploides, (n), y de polaridades (sexos) opuestos, además se producen en estructuras especiales, los gametangios.
Existen varios tipos de reproducción sexual:
- Isogamia: unión de gametos de igual forma y tamaño pero de polaridades distintas.
- Anisogamia: Unión de gametos distintos en forma y tamaño y de polaridad opuesta.
- Oogamia: es un caso especial de anisogamia pero con un gameto femenino inmóvil y de mayor tamaño que el masculino.
Los gametos, al igual que las esporas, reciben distintos nombres. Los gametos femeninos se llaman ovocélula, oosfera, óvulo; y los masculinos anterozoide, anterozoo, espermatozoide, espermatozoo, espermacio que puede ser inmóvil en algunos hongos.
Los gametangios también reciben nombres especiales, existen los mismos tipos que los de los esporangios. Los gametangios primitivos son unicelulares y pueden estar recubiertos por una o varias paredes celulares.
Los gametangios más evolucionados son los pluricelulares con cubierta de protección. En la oogamia, el gametangio masculino se llama anteridio y posee un menor tamaño y forma filiforme, el femenino se llama oogonio y tiene mayor tamaño y forma esférica. De Briófitos en adelante, al masculino se le llama anteridio y al femenino arquegonio.
Existen casos especiales de gametangiogamia en la que la fecundación se lleva a cabo por fusión de gametangios y posterior unión de gametos; de somatogamia en la que no se producen gametangios y se copulan células somáticas; y apomixis, reproducción sexual sin fecundación, las células somáticas hacen el papel de germinales. En este artículo puedes encontrar más información sobre cómo se forman los espermatozoides de las plantas.
La reproducción de las plantas angiospermas
La reproducción sexual de las plantas angiospermas es el ejemplo más popular. Es la reproducción de las plantas con flores. Cuando un grano de polen llega al pistilo de la flor, se produce la polinización. Al entrar en su interior, se produce la generación de las células germinales o gametos masculinos que fecundan los óvulos que hay en el gineceo mediante el tubo polinizador.
Una vez los óvulos están fecundados, proceso conocido como fertilización, se forman las semillas, y una vez formadas éstas, el gineceo se convierte en el fruto que habitualmente conocemos como resultado de la reproducción de las plantas.
En la fecundación se produce la unión dos células haploides con distinto material genético, que se unen formando un individuo de genotipo diferente del de los padres y por este motivo se llama reproducción sexual.
Origen y evolución de la sexualidad
Se piensa que los eucariotas surgieron hace unos 600 M .A. a partir de procariotas debido a la invasión de nuevos hábitats y que estos eucariotas primitivos son los protoctistas.
La aparición de la reproducción sexual se produjo a partir del perfeccionamiento de la mitosis y la aparición de la meiosis y ciclo sexual. El origen de la mitosis pudo producirse en los protoctistas y especialmente en los dinoflagelados ya que su mitosis es “anómala”, no hay huso mitótico.
El origen de la reproducción sexual también pudo haberse dado en protoctistas al mismo tiempo que la mitosis debido a que estos organismos poseen ambos procesos incompletos. Surgió como consecuencia de la ventaja adaptativa que suponía ya que al haber recombinación cromosómica es exploradora e innovadora y facilita la colonización de nuevos hábitats. En la reproduccion de las plantas se da este tipo como ya hemos visto junto con la reproduccion asexual.
Si quieres ampliar más información sobre la reproducción sexual y asexual de las plantas o la botánica en general, te recomendamos el libro Strasburger: Tratado de botánica del que se ha extraído parte de esta información. Además, es el libro clásico y de cabecera de cualquier estudiante de botánica.
SEMANA DEL 5 AL 8 DE JULIO
TIPOS DE REPRODUCCIÓN ASEXUAL
La reproducción asexual o asexualidad involucra un tipo de reproducción donde la constitución genética de los dos organismos que se producen es idéntica a la del progenitor.
Existen distintos tipos de reproducción asexual entre los que se encuentran la bipartición o simple fisión, la gemación, la fragmentación y la esporulación.
• La bipartición o fisión, es un tipo de reproducción asexual en el que la célula se divide dando origen a dos células genéticamente idénticas entre sí pero de menor tamaño que la inicial. Esta modalidad de reproducción se da principalmente en organismos unicelulares como los protozoos y las bacterias.
En las bacterias, el proceso se denomina fisión binaria transversal, y se caracteriza porque el material hereditario (DNA) de la bacteria, se duplica y luego se separa; y la pared celular se forma en el interior y en sentido transversal.
• La gemación ocurre en organismos unicelulares y multicelulares. En la superficie del progenitor se forma una gema que crece y se estrangula hasta separarse por completo del organismo original. Las células producidas pueden tener vida propia o formar colonias si permanecen unidas al organismo parental. La levadura es un hongo que se reproduce por gemación. La gemación también ocurre en organismos pluricelulares. Un ejemplo de esto lo constituye la hidra. Sobre la superficie corporal de este organismo pluricelular, aparecen abultamien-tos que luego darán origen a nuevas hidras, genéticamente idénticas a su progenitor, pero de menor tamaño.
• La fragmentación. En este tipo de reproducción asexual se origina un nuevo organismo a partir de fragmentos del organismo progenitor. Ésta se da en organismos como las plenarias, que cuando se cortan por la mitad originan un nuevo organismo a partir de cada mitad.
A medida que se asciende en la escala zoológica, esta modalidad de reproducción va desapareciendo, y es reemplazada por mecanismos de regeneración de tejidos dañados. Por ejemplo, una lagartija (Leo-laemus montícola) puede regenerar parte de su cola cuando la ha perdido, la estrella de mar regenera parte del brazo perdido; sin embargo, el hombre tiene menos facilidad para regenerar estructuras dañadas: el crecimiento de uñas, del cabello y la cicatrización son algunos ejemplos.
SEMANA DEL 23 AL 27 DE MAYO
Repaso de genética
1. Se cruzan dos plantas de flores color naranja y se obtiene una descendencia formada por 30 plantas
de flores rojas, 60 de flores naranja y 30 de flores amarillas. ¿Qué descendencia se obtendrá al
cruzar las plantas de flores naranjas obtenidas, con las rojas y con las amarillas también obtenidas?
Razona los tres cruzamientos.
2. Una planta de jardín presenta dos variedades: una de flores rojas y hojas alargadas y otra de flores
blancas y hojas pequeñas. El carácter color de las flores sigue una herencia intermedia, y el carácter
tamaño de la hoja presenta dominancia del carácter alargado. Si se cruzan ambas variedades,
¿Qué proporciones genotípicas y fenotípicas aparecerán en la F2? ¿Qué proporción de las flores rojas
y hojas alargadas de la F2 serán homocigóticas?
3. Indica el genotipo de un hombre calvo cuyo padre no era calvo, el de su esposa que no es calva,
pero cuya madre sí lo era, y el de sus futuros hijos.
4. Se cruzan tomates rojos híbridos y de tamaño normal homocigóticos con la variedad amarilla enana.
¿Qué proporción de los tomates rojos que salen en la F2 serán enanos? (Los alelos dominantes
son color rojo y tamaño normal).
5. Un perro de pelo negro, cuyo padre era de pelo blanco, se cruza con una perra de pelo gris, cuya
madre era negra. Sabiendo que el pelaje negro domina sobre el blanco en los machos, y que en las
hembras negro y blanco presentan herencia intermedia, explica cómo serán los genotipos de los
perros que se cruzan y qué tipos de hijos pueden tener respecto del carácter considerado.
SEMANA DEL 16 AL 20 DE MAYO
Lección 21: Codominancia o dominancia incompleta
También se denomina herencia intermedia y se caracteriza porque el heterocigoto presenta un fenotipo intermedio al que producen los individuos homocigóticos; es decir el heterocigoto no manifiesta la misma relación fenotípica del homocigoto dominante, como ocurre en la herencia de tipo Mendeliano. Cuando hay dominancia incompleta entre dos alelos las proporciones fenotípicas en la F2 son de 1:2:1 y el fenotipo describe el genotipo; diferente a la proporción Mendeliana clásica 3:1.
Para este caso y para el análisis de los problemas que tienen que ver con este principio se emplea una simbología que consiste en utilizar una letra base en mayúscula, igual para los genes y un superíndice que puede ser una letra, un número o un símbolo que indica la variabilidad del gen, para esa misma característica por ejemplo:
Fenotipo color rojo : FRFR
Fenotipo color blanco : FrFr
Fenotipo color rosado : FRFr
Ejemplo
El color del pelaje en el ganado Shorthorn en el que se presentan tres tipos de color: rojo, blanco y ruano; en donde el ruano se produce al aparearse un animal rojo con uno blanco.

SEMANA DEL 2 AL 6 DE MAYO
Leyes De Mendel
Las leyes de Mendel fueron desarrolladas por un científico genetista, considerado como el padre de la genética: Gregor Mendel. De allí su nombre. Este científico realizo experimentos que permitieron dilucidar elementos fundamentales de la herencia genética, como con un ejemplo de ley de Mendel, donde se explican los rasgos descendientes que se pueden predecir a través de las características de los progenitores de una especie, desde animales, plantas y hasta seres humanos.
Este científico fue quien acuñó algunos de los términos más conocidos de la genética, como son los términos “dominante” y “recesivo”, que son factores de la herencia presentes en las características y rasgos hereditarios en los organismos, todo esto a través de las tres Leyes de Mendel.
Fue decisivo el Ensayo Sobre Los Híbridos Vegetales que realizó en 1866, donde finalmente se formulaban las 3 Leyes De Mendel que fueron nombradas ante su apellido. Y que estaban compuestas por cruces inter-especies y experimentos que fueron llevados a un análisis estadístico. Sin embargo, estos estudios no fueron tomados en cuenta hasta mucho después de ser publicados, en el año 1900. A continuación te damos una introducción a las Leyes de Gregor Mendel:
Fue decisivo el Ensayo Sobre Los Híbridos Vegetales que realizó en 1866, donde finalmente se formulaban las 3 Leyes De Mendel que fueron nombradas ante su apellido. Y que estaban compuestas por cruces inter-especies y experimentos que fueron llevados a un análisis estadístico. Sin embargo, estos estudios no fueron tomados en cuenta hasta mucho después de ser publicados, en el año 1900. A continuación te damos una introducción a las Leyes de Gregor Mendel:
Ley De Mendel
La primera ley de Mendel, también llamada: Ley de la uniformidad de los híbridos de la primera generación, o simplemente Ley de la Uniformidad. Esta ley dicta que, al cruzar dos variedades de una especie de raza pura, cada uno de los híbridos de la primera generación tendrá caracteres determinados similares en su fenotipo. Esto se debe a que las razas puras tienen un gen dominante o un gen recesivo. El genotipo dominante será entonces el que determine la característica o características principales de la primera generación del cruce, pero al mismo tiempo, también serán similares fenotípicamente entre sí, es decir, entre cada individuo de la primera generación.
En el experimento realizado por Mendel para obtener la primera de las leyes de Mendel, utilizaba una especie de chícharos que producían semillas amarillas como gen dominante y otra que tenía un gen recesivo que producía semillas verdes, por lo tanto, el alelo que llamaremos “A” daba el color amarillo por encima del alelo “a” que producía el color verde. El producto del cruce eran plantas que producían semillas amarillas. Sigue leyendo aquí para aprender más sobre la Ley De Mendel.
Segunda Ley De Mendel
La segunda ley de Mendel, también conocida como la Ley de la Segregación, Ley de la Separación Equitativa, o hasta Ley de Disyunción de los Alelos. Esta dictamina que para que exista la reproducción de dos individuos de una especie, primero debe existir la separación del alelo de cada uno de los pares para que de esta manera se transfiera la información genética al hijo. Un alelo es, la variante genética que permite determinar un rasgo o carácter. Existen entonces, alelos dominantes y alelos recesivos.
Por esto, es que la segunda de las leyes de Mendel se la llama como de segregación o separación, ya que cada padre, aporta un alelo que se separa de cada uno, para formar un individuo en una nueva generación. Sigue leyendo aquí para aprender más sobre la Segunda Ley De Mendel. Mendel, en su experimento, obtuvo solo semillas amarillas en la primera generación, pero en la segunda generación, los alelos se separaron para formar nuevas semillas verdes en menor proporción que las amarillas, pero aun así existentes. Esta sería la proporción:
Tercera Ley De Mendel
La tercera ley de Mendel, también llamada Ley de la Herencia Independiente de Caracteres o Ley de la Asociación Independiente. Según Mendel, hay rasgos heredados que se obtienen de forma independiente, sin relación con el fenotipo, lo cual no afecta al patrón de herencia de otros rasgos. Esta ley se cumple en los genes que no están ligados, es decir que se encuentran en diferentes cromosomas o que están en zonas muy separadas del mismo cromosoma.
Mendel, para concluir la tercera de las leyes de Mendel, realizó un cruce de plantas de chícharos que producían semillas amarillas y llanas, con chícharos que producían semillas verdes y con textura irregular. Estas eran homocigóticas para los dos caracteres de textura y color. Se concluía que la ley de uniformidad estaba presente, pues con la primera generación se pudo obtener semillas amarillas y lisas.
Sin embargo, al cruzar esta primera generación para obtener una segunda generación, se observan nuevos tipos de semillas con caracteres diversos pero relacionados con la generación parental, se obtuvieron semillas amarillas y lisas, amarillas y rugosas, verdes y lisas, y verdes y rugosas. Sigue leyendo aquí para aprender más acerca de Tercera Ley De Mendel.
Leyes De Mendel Resumidas
Para terminar con las Leyes De Mendel Resumidas, se puede decir que, la primera Ley de Mendel dice que si se cruzan dos padres de raza pura con diferentes rasgos, la primera generación tendrá similitudes entre sí y guardará un carácter del padre con el alelo dominante. La segunda ley dice que, los factores genéticos se separan de cada uno de los padres en alelos individuales que se juntarán para procrear una descendencia con las características de la primera generación, pero en la segunda generación, se manifiestan nuevos rasgos genéticos observados en los padres pero unidos de manera aleatoria en la descendencia de la primera generación. Y la tercera ley de Mendel dice que, además existen rasgos generados de forma independiente, a través de cromosomas alejados que no intervienen entre sí, y al igual que en la segunda ley, esta tercera de las leyes de Mendel se manifiesta con más claridad en la segunda generación de individuos.
LOS SIGUIENTES ESTUDIANTES DEBEN PRESENTAR UN EXAMEN COMO NIVELACIÓN DEL PRIMER PERIODO EN BIOLOGÍA
JOSE MARÍA CÓRDOBA
ELENA BORSERINI
JULIETA UMAÑA
CARLOS ESCOBAR
JUAN DIEGO PINEDA
JOSE CARLOS VELÁSQUEZ
TEMAS RELEVANTES PARA LA NIVELACIÓN
ESPECIACION ALOPATRICA Y SIMPATRICA
CARACTERES HOMÓLOGOS Y ANÁLOGOS
SELECCIÓN SEXUAL
EFECTO FUNDADOR Y CUELLO DE BOTELLA
EVIDENCIAS DIRECTAS E INDIRECTAS DE LA EVOLUCIÓN
TEORIA DE DARWIN
EQUILIBRIO DE HARDY WEINBERG
SEMANA DEL 14 AL 18 DE MARZO
EQUILIBRIO DE HARDY-WEINBERG
La ley de Hardy-Weinberg establece que en una población suficientemente grande, en la que los apareamientos se producen al azar y que no se encuentra sometida a mutación, selección o migración, las freecuencias génicas y genotípcas se mantienen constantes de una generación a otra, una vez alcanzado un estado de equilibrio que en loci autosómicos se alcanza tras una generación.
Se dice que una población está en equilibrio cuando los alelos de los sistemas polimórficos mantienen su frecuencia en la población a través de las generaciones.
Para lograr el equilibrio genético, según el matemático inglés Hardy y el médico alemán Weinberg, se deben dar varias condiciones:
Se dice que una población está en equilibrio cuando los alelos de los sistemas polimórficos mantienen su frecuencia en la población a través de las generaciones.
Para lograr el equilibrio genético, según el matemático inglés Hardy y el médico alemán Weinberg, se deben dar varias condiciones:
- La población debe ser infinitamente grande y los apareamientos al azar (panmícticos).
- No debe existir selección, es decir, cada genotipo bajo consideración debe poder sobrevivir tan bien como cualquier otro (no hay mortalidad diferencial) y cada genotipo debe ser igualmente eficiente en la producción de progenie (no hay reproducción diferencial).
- No debe existir flujo génico, es decir, debe tratarse de una población cerrada donde no haya inmigración ni emigración.
- No debe haber mutaciones, a excepción que la mutación se produzca en sentido inverso con frecuencias equivalentes, por ejemplo, A muta hacia A' con la misma frecuencia con la que A' muta hacia A.
Toda demostración de la ley de Hardy-Weinberg implica el principio básico de la teoría de la probabilidad, esto es, que la probabilidad de ocurrencia simultánea de dos o más eventos independientes es igual al producto de las probabilidades de cada evento. Normalmente, la frecuencia de cada alelo representa su probabilidad de ocurrencia. De modo que para obtener la probabilidad de un genotipo dado en la progenie, se multiplican las frecuencias de los alelos involucrados entre sí.
Dadas las frecuencias génicas (alélicas) en el pool génico de una población, es posible calcular (con base en la probabilidad de la unión de gametos) las frecuencias esperadas de los genotipos y fenotipos de la progenie. Si p = porcentaje del alelo A (dominante) y q = el porcentaje del alelo a (recesivo), se puede utilizar el método del damero para producir todas las posibles combinaciones al azar de estos gametos.
Dadas las frecuencias génicas (alélicas) en el pool génico de una población, es posible calcular (con base en la probabilidad de la unión de gametos) las frecuencias esperadas de los genotipos y fenotipos de la progenie. Si p = porcentaje del alelo A (dominante) y q = el porcentaje del alelo a (recesivo), se puede utilizar el método del damero para producir todas las posibles combinaciones al azar de estos gametos.
| Hembras | |||
| Gametos | p (A) | q (a) | |
| Machos | p (A) | p2 (AA) | pq (Aa) |
| q (a) | pq (Aa) | q2 (aa) | |
Obsérvese que p + q = 100% (o 1), es decir, los porcentajes de los gametos A y a deben igualar al 100%) para incluir a todos los gametos del pool génico. De este modo, si solo conozco la frecuencia de un alelo y sé que el sistema genético analizado solo tiene 2, puedo calcular la frecuencia del segundo. Más fácilmente, puedo expresar p y q como proporciones, igualando p + q = 1.
Las frecuencias genotípicas (cigóticas) en la siguiente generación sumadas también deben igualar 1 o 100%, y pueden resumirse como sigue:
(p + q)2 = p2 + 2pq + q2 = 1
AA Aa aa
Así, dado que los gametos serán p o q, y que cada individuo recibe dos gametos, la siguiente generación se espera que sea: p2 la fracción de homocigóticos dominantes (AA), 2pq, la fracción de los heterocigóticos (Aa) y q2, la fracción de los homocigóticos recesivos (aa).
En pocas palabras, en ausencia de factores disruptivos (mutación, flujo génico desbalanceado, selección natural y deriva génica), las frecuencias alélicas y genotípicas en un locus de una población diploide panmíctica se repetirán de generación en generación. Es importante destacar que en caso de darse una alteración de las frecuencias, éstas restablecerán el equilibrio tras una generación de cruzamientos al azar.
Por ejemplo, supongamos una población hipotética con las frecuencias genotípicas:
En pocas palabras, en ausencia de factores disruptivos (mutación, flujo génico desbalanceado, selección natural y deriva génica), las frecuencias alélicas y genotípicas en un locus de una población diploide panmíctica se repetirán de generación en generación. Es importante destacar que en caso de darse una alteración de las frecuencias, éstas restablecerán el equilibrio tras una generación de cruzamientos al azar.
Por ejemplo, supongamos una población hipotética con las frecuencias genotípicas:
D(AA) = 0.10Las frecuencias génicas se calculan:
H(Aa) = 0.20
R(aa) = 0.70
p(A) = D + 1/2 H = 0.10 + 0.10 = 0.20Las frecuencias esperadas de acuerdo a la ley de Hardy-Weinberg son:
q(a) = 1/2H + R = 0.10 + 0.70 = 0.80
p2[AA] = 0.04
2pq[Aa] = 0.32
q2[aa] = 0.64
SEMANA DEL 7 DE MARZO AL 11 DE MARZO
RESUELVE LA SIGUIENTE ACTIVIDAD
COPIA LA IMAGEN E IMPRIME PARA RESOLVERLA Y LLÉVALA PEGADA EN EL
CUADERNO


SEMANA DEL 29 DE FEBRERO AL 4 DE MARZO
Los cuellos de botella y el efecto fundador
La deriva genética puede causar grandes pérdidas de variabilidad genética en las poblaciones pequeñas.
Cuando el tamaño de una población se reduce durante al menos una generación, se porduce un cuello de botella poblacional. Debido a que la deriva genética actúa más rápidamente reduciendo la variabilidad genética en las poblaciones pequeñas, pasar por un cuello de botella puede hacer que disminuya mucho la variabilidad genética de una población, incluso aunque el cuello de botella no dure muchas generaciones. Las bolsas de canicas de más abajo ejemplifican este proceso, donde, en la generación 2, una extracción singularmente pequeña crea un cuello de botella.
 |
Una disminución de la variabilidad genética implica que la población puede no adaptarse a las nuevas presiones selectivas, como el cambio climático o un cambio en los recursos disponibles, debido a que la variabilidad genética sobre la que actuaría la selección puede haber desaparecido ya de la población.
 |
Un ejemplo de cuello de botella
Los elefantes marinos del norte tienen poca variabilidad genética, probablemente, por un cuello de botella por el que les hicieron pasar los hombres en la década de 1890. La caza hizo disminuir el censo poblacional hasta tan sólo 20 individuos a finales del siglo XIX. Desde entonces, su población ha «rebotado» hasta más de 30 000, pero sus genes todavía llevan las señales de ese cuello de botella: su variabilidad genética es mucho menor que la de una población de elefantes marinos del sur que no sufrió una caza tan intensa.
Los elefantes marinos del norte tienen poca variabilidad genética, probablemente, por un cuello de botella por el que les hicieron pasar los hombres en la década de 1890. La caza hizo disminuir el censo poblacional hasta tan sólo 20 individuos a finales del siglo XIX. Desde entonces, su población ha «rebotado» hasta más de 30 000, pero sus genes todavía llevan las señales de ese cuello de botella: su variabilidad genética es mucho menor que la de una población de elefantes marinos del sur que no sufrió una caza tan intensa.
El efecto fundador
El efecto fundador se produce cuando unos pocos miembros de la población original establecen una colonia. Este pequeño tamaño poblacional implica que la colonia puede tener:
El efecto fundador se produce cuando unos pocos miembros de la población original establecen una colonia. Este pequeño tamaño poblacional implica que la colonia puede tener:
- poca variabilidad genética, respecto a la población original.
- una muestra no aleatoria de los genes de la población original.
Por ejemplo, la población afrikáner de colonos holandeses de Sudáfrica desciende principalmente de unos pocos colonos. En la actualidad, en la población afrikáner existe una frecuencia excepcionalmente alta del gen que causa la corea de Huntington debido a que dio la casualidad de que aquellos colonos holandeses originales eran portadores de ese gen con una frecuencia excepcionalmente alta. Este efecto es fácil de reconocer en las enfermedades genéticas pero, por supuesto, los eventos fundadores afectan a la frecuencia de todo tipo de genes
Semana del 22 al 26 de febrero
Los mecanismos de la microevolución
Los cambios microevolutivos se producen de unas pocas formas básicas. Los procesos que pueden afectar directamente las frecuencias génicas de una población son la mutación, migración, deriva genética y selección natural.
Imagina que en una población de escarabajos observas un aumento de la frecuencia de los genes para el color marrón y una disminución de la frecuencia de los genes para el color verde. Cualquier combinación de los mecanismos de la microevolución puede ser responsable de este patrón, por lo que parte del trabajo de los científicos es averiguar cuál de estos mecanismos fue el causante del cambio:
Mutación
Algunos genes «verde» mutaron aleatoriamente, transformándose en genes «marrón» (aunque, dado que cualquier mutación en especial es rara, este proceso por sí solo no puede ser responsable de un gran cambio en la frecuencia alélica en una generación).

Migración (o flujo génico)
Algunos escarabajos con genes marrón inmigraron desde otra población, o algunos escarabajos que tenían genes verde emigraron.

Deriva genética
Al reproducirse los escarabajos acabaron en la descendencia más genes marrón que genes verde, simplemente por el azar. En el diagrama de la derecha, los genes marrón están presentes con una frecuencia ligeramente superior en la descendencia (29%) que en la generación parental (25%).

Selección natural
Los escarabajos con genes marrón evitaron ser cazados y sobrevivieron para reproducirse con más frecuencia que los escarabajos con genes verde, por lo que pasaron más genes marrón a la siguiente generación.
 |
:::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::
Existen varias pruebas de evolución en todas las especies, estas pueden ser directas o indirectas.
Entre las evidencias directas se cuentan los fósiles de animales, como huesos, excrementos petrificados, impresiones de piel o huellas y huevos petrificados, estos pueden evidenciar cambios físicos que han sufrido las especies, así se pueden determinar los antepasados de estas.
Las evidencias indirectas demuestran el parentesco entre especies, ya sea comparando algunos órganos que tienen funciones o estructura parecida, o el análisis de órganos vestigiales (anatomía comparada) comparando el desarrollo embrionario (se ha encontrado que muchos animales tienen la misma forma en las primeras semanas). Otra evidencia indirecta es a bioquímica comparada, en la que se analizan muestras de sangre de diferentes animales comparando las moléculas de hemoglobina para ver sus secuencias de ADN y comparar sus similitudes.
Semana del 8 al 12
LA SELECCIÓN NATURAL
LAS MUTACIONES
Una mutación es un cambio en el ADN, el material hereditario de los seres vivos. El ADN de un organismo influye en su aspecto físico, en su comportamiento y en su fisiología — en todos los aspectos de su vida. Por lo tanto, un cambio en el ADN de un organismo puede producir cambios en todos los aspectos de su vida.
Las mutaciones son aleatorias
Las mutaciones pueden ser beneficiosas, neutras o dañinas para el organismo, pero las mutaciones no «intentan» proporcionar lo que el organismo «necesita». En este sentido, las mutaciones son aleatorias — el hecho de que una mutación concreta suceda o no, no está relacionado con lo útil que sería.
No todas las mutaciones son relevantes para la evolución
Dado que todas las células de nuestro cuerpo contienen ADN, hay multitud de lugares en los que pueden producirse las mutaciones; sin embargo, no todas las mutaciones son relevantes para la evolución. Las mutaciones somáticas son las que se producen en las células no reproductoras y no se transmiten a la descendencia.
Por ejemplo, el color amarillo de la mitad de esta manzana Red Delicious fue causado por una mutación somática, por lo que sus las semillas no son portadoras de la mutación.

::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::
2016
El código genético
Actividad para el 10 de febrero
ve al siguiente link, observa el vídeo y sigue las flechas para que al final puedas resolver la pregunta en el cuaderno. La cual se sustentara en la clase del 10 de febrero
Link: Contenido material genetico
o copia y pega el siguiente link en la barra de dirección
http://contenidosparaaprender.mineducacion.gov.co/G_8/S/S_G08_U02_L07/S/S_G08_U02_L07/S_G08_U02_L07_03_06.html
::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::
Hola chicos
actividad para el 9 de febrero
A continuación dejo el link donde deben realizar la actividad que se indica, al finalizar la actividad que se propone realiza el ejercicio que aparece en el cuaderno
Se revisara la próxima clase
2016
:::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::
ESTUDIANTES QUE DEBEN PRESENTAR NIVELACIONES EN LA ASIGNATURA DE BIOLOGÍA PARA CUARTO PERIODO
BIOLOGÍA: FECHA DE LA NIVELACIÓN 19 DE NOVIEMBRE
RODRIGO ARDILA: presentar cuaderno al día, con todas las notas pendientes, de las cuales se harán preguntas orales
SEMANA DEL 28 DE SEPTIEMBRE AL 2 DE OCTUBRE
LOS ECOSISTEMAS

El ecosistema es el conjunto de especies de un área
determinada que interactúan entre ellas y con su ambiente abiótico; mediante
procesos como la depredación, el parasitismo, la competencia y la simbiosis, y
con su ambiente al desintegrarse y volver a ser parte del ciclo de energía y de
nutrientes. Las especies del ecosistema, incluyendo bacterias, hongos, plantas
y animales dependen unas de otras. Las relaciones entre las especies y su
medio, resultan en el flujo de materia y energía del ecosistema.
El significado del concepto de ecosistema ha evolucionado
desde su origen. El término acuñado en los años 1930s, se adscribe a los
botánicos ingleses Roy Clapham (1904-1990) y Sir Arthur Tansley (1871-1955). En
un principio se aplicó a unidades de diversas escalas espaciales, desde un
pedazo de tronco degradado, un charco, una región o la biosfera entera del
planeta, siempre y cuando en ellas pudieran existir organismos, ambiente físico
e interacciones.
Más recientemente, se le ha dado un énfasis geográfico y se
ha hecho análogo a las formaciones o tipos de vegetación; por ejemplo,
matorral, bosque de pinos, pastizal, etc. Esta simplificación ignora el hecho
de que los límites de algunos tipos de vegetación son discretos, mientras que
los límites de los ecosistemas no lo son. A las zonas de transición entre
ecosistemas se les conoce como “ecotonos”.
A grandes rasgos se habla de tres tipos de ecosistemas:
a. Acuático:
Esta clase de ecosistema los seres vivos se desarrollan en el agua. Estos, adquieren características físicas muy similares entre sí como consecuencia de su adaptación al agua. En este ecosistema las variaciones de temperaturas no son muy marcadas, por lo que esta no afecta la supervivencia de los seres vivos. Este ecosistema es el de mayor tamaño ya que representan el 75%. Dentro de los ecosistemas acuáticos se encuentran los siguientes:
Bentónico: estos se ubican en el fondo de los ecosistemas acuáticos. En aquellos que no son muy profundos, los principales habitantes son algas. En los de mayor profundidad, la mayoría son consumidores.
Nectónicos: estos animales se desplazan con total libertad ya que gracias a sus medios de locomoción pueden adaptarse a las corrientes de agua.
Plactónicos: estos seres vivos viven flotando en el agua terrestre o marina y son arrastrados por las corrientes de agua, no se trasladan por movimientos propios.
Neustónicos: estos viven sobre la superficie del agua, flotando.
b. Aéreo:
Este tipo de ecosistema tiene la particularidad de ser de transición. Ningún ser vivo lo habita permanentemente, sino que tienen que descender a la tierra para el descanso, alimentación o procreación, por lo que no resulta autosuficiente. A causa de esto, algunos lo ubican dentro del ecosistema terrestre.
Terrestre
Este ecosistema se desarrolla sobre la superficie de la Tierra llamada Biósfera. Los individuos más numerosos en este ecosistema son los insectos, de los que existen 900.000 especies. Las aves ocuparían el segundo lugar, con unas 8.500 especies. En tercer lugar, los mamíferos de los que hay 4.100 especies. A diferencia del ecosistema acuático, en el terrestre los individuos presentan características mucho más variadas, esto se debe a los numerosos factores que condicionan a las especies. Entre estos los más importantes son: la radiación solar, la disponibilidad de agua, nutrientes y luz. Otra característica de este ecosistema es la necesidad que tienen, tanto los vegetales como animales, de agua para la hidratación de sus organismos, por lo que sin ella no podrían subsistir.
SEMANA DEL 21 AL 25 DE SEPTIEMBRE
HOLA CHICOS, AQUÍ LES DEJO EL LINK DE LA INFOGRAFIA OBSERVADA EN CLASES.
http://www.educ.ar/recursos/ver?id=20073
Niveles de Organización

La biología se ocupa de analizar jerarquías o niveles de organización que van desde la célula a los ecosistemas. Este concepto implica que en el universo existen diversos niveles de complejidad.Por lo tanto es posible estudiar biología a muchos niveles, desde un conjunto de organismos (comunidades) hasta la manera en que funciona una célula o la función de las moléculas de la misma.
En orden decreciente mencionaremos los principales niveles de organización:
| Biosfera: La suma de todos los seres vivos tomados en conjunto con su medio ambiente. En esencia, el lugar donde ocurre la vida, desde las alturas de nuestra atmósfera hasta el fondo de los océanos o hasta los primeros metros de la superficie del suelo (o digamos mejor kilómetros sí consideramos a las bacterias que se pueden encontrar hasta una profundidad de cerca de 4 Km. de la superficie). Dividimos a la Tierra en atmósfera (aire),litosfera (tierra firme), hidrosfera (agua), y biosfera (vida). | |
| Ecosistema: La relación entre un grupo de organismos entre sí y su medio ambiente. Los científicos a menudo hablan de la interrelación entre los organismos vivos. Dado, que de acuerdo a la teoría de Darwin los organismos se adaptan a su medio ambiente, también deben adaptarse a los otros organismos de ese ambiente. | |
| Comunidad: Es la relación entre grupos de diferentes especies. Por ejemplo, las comunidades del desierto pueden consistir en conejos, coyotes, víboras, ratones, aves y plantas como los cactus. La estructura de una comunidad puede ser alterada por cosas tales como el fuego, la actividad humana y la sobrepoblación. | |
| Especie: Grupo de individuos similares que tienden a aparearse entre sí dando origen a una cría fértil. Muchas veces encontramos especies descriptas, no por su reproducción (especies biológicas) sino por su forma (especies anatómicas). | |
| Poblaciones: Grupos de individuos similares que tienden a aparearse entre sí en un área geográfica limitada. Esto puede ser tan sencillo como un campo con flores separado de otro campo por una colina sin flores. | |
| Individuo: Una o más células caracterizadas por un único tipo de información codificada en su ADN. Puede ser unicelular o multicelular. Los individuos multicelulares muestran tipos celulares especializados y división de funciones en tejidos, órganos y sistemas. | |
| Sistema: (en organismos multicelulares). Grupo de células, tejidos y órganos que están organizados para realizar una determinada función, p.ej. el sistema circulatorio. | |
| Órganos: (en organismos multicelulares). Grupo de células o tejidos que realizan una determinada función. Por ejemplo el corazón, es un órgano que bombea la sangre en el sistema circulatorio. | |
| Tejido: (en organismos multicelulares). Un grupo de células que realizan una determinada función. Por ejemplo el tejido muscular cardíaco. | |
| Célula: la más pequeña unidad estructural de los seres vivos capaz de funcionar independientemente. Cada célula tiene un soporte químico para la herencia (ADN), un sistema químico para adquirir energía etc. | |
| Organela: una subunidad de la célula. Una organela se encuentra relacionada con una determinada función celular p.ej. la mitocondria (el sitio principal de generación de ATP en eucariotas). | |
Moléculas, átomos, y partículas subatómicas: los niveles funcionales fundamentales de la bioquímica.
|
 |
 |
SEMANA DEL 14 AL 18 DE SEPTIEMBRE
| LOS CICLOS BIOGEOQUÍMICOS |  |
El concepto de ciclo biogeoquímico se usa para describir la distribución y transporte de materiales, los cuales controlan el recambio y transformación de éstos en los ambientes terrestres, acuáticos y atmosféricos. Los ciclos biogeoquímicos constituyen un sistema regulador de la hidrosfera y la biosfera. Estos ciclos describen los movimientos y las interacciones de los elementos químicos esenciales para la vida a través de la geosfera y la geosfera, a través de procesos físicos, químicos y biológicos. Los flujos de los elementos pueden ser abiertos, como el flujo de energía o cerrados, como el ciclo de la materia.
El ciclo de la materia es una interacción permanente entre la fase biótica y la fase abiótica, es un proceso sin principio ni fin; es decir, un reciclaje combinado y continuo, en una serie de procesos autorregulados; los deshechos son el punto de partida para formar algo nuevo.
Los principales elementos químicos son: carbono, hidrógeno, nitrógeno, oxígeno, fósforo, azufre o los contaminantes, los ciclos de estos elementos se combinan de diferentes maneras e interrelacionan entre sí. Un solo elemento puede convertirse en el factor limitante en el desarrollo de un ecosistema. Por ejemplo la oferta de nitrógeno puede limitar los procesos vitales en los océanos. La comprensión de los ciclos biogeoquímicos es esencial para entender el funcionamiento de la tierra como sistema.
Los ciclos se usan para medir la dinámica del recambio comparando las magnitudes en el depósito y los flujos en diferentes compartimentos del ecosistema. De particular interés son las escalas espaciales y temporales de las transformaciones y las fases de transición.
Los aspectos básicos de los ciclos biogeoquímicos son:
- La distribución de materiales- localización y tamaño del depósito.
- El transporte- patrones y ratas de flujo.
- La transformación- rata de flujo del depósito a otro componente.
- Tiempo de residencia- Tiempo de almacenamiento.
Los ciclos biogeoquímicos generalmente se conceptualizan en modelos de compartimentos y se visualizan convenientemente por medio de cuadros y flechas (fig. 45) Entre los 80 elementos que se encuentran en el suelo, sólo una tercera parte son componentes esenciales en plantas y animales. Entre los elementos principales que constituyen la materia orgánica tenemos: C, H, O, N, P, S, mientras que otros cumplen la función de matrices iónicas o estructuras de soporte: Ca, Mg, Si, K, Na, Cl, F). Los metales esenciales traza, se encuentran generalmente como coenzimas (Fe, Mn, Co, Cu, Zn, Se, Mo). Hay algunos elementos importantes que no se usan por los organismos: uno muy abundante es el aluminio y otros que son muy tóxicos como: Hg>Cd>Pb. La composición en elementos de la biomasa, el agua de mar y la corteza terrestre se resume en la tabla 4.
SEMANA DEL 7 AL 11 DE SEPTIEMBRE
La fotosíntesis es el proceso de elaboración de los alimentos por parte de las plantas. Los árboles y las plantas usan la fotosíntesis para alimentarse, crecer y desarrollarse.
Para realizar la fotosíntesis, las plantas necesitan de la clorofila, que es una sustancia de color verde que tienen en las hojas. Es la encargada de absorber la luz adecuada para realizar este proceso. A su vez, la clorofila es responsable del característico color verde de las plantas.

Foto: EFE
Para realizar la fotosíntesis, las plantas necesitan de la clorofila, que es una sustancia de color verde que tienen en las hojas.
El proceso completo de la alimentación de las plantas consiste básicamente en:
a- Absorción: Las raíces de las plantas crecen hacia donde hay agua. Las raíces absorben el agua y los minerales de la tierra.
b- Circulación: Con el agua y los minerales absorbidos por las raíces hasta las hojas a través del tallo.
c- Fotosíntesis: Se realiza en las hojas, que se orientan hacia la luz. La clorofila de las hojas atrapa la luz del Sol. A partir de la luz del Sol y el dióxido de carbono, se transforma la savia bruta en savia elaborada, que constituye el alimento de la planta. Además la planta produce oxígeno que es expulsado por las hojas.
d- Respiración: Las plantas , al igual que los animales, tomando oxígeno y expulsando dióxido de carbono. El proceso se produce sobre todo en las hojas y el los tallos verdes. La respiración la hacen tanto de día como por la noche, en la que, ante la falta de luz, las plantas realizan solamente la función de respiración.
La importancia de la fotosíntesis
La fotosíntesis hace que las plantas generen oxígeno, que es el elemento que respiran todos los seres vivos. Además, las plantas consumen gases tóxicos, como el dióxido de carbono.
SEMANA DEL 31 DE AGOSTO AL 4 DE SEPTIEMBRE
Cadenas alimentarias o tróficas |
En los ecosistemas se establecen relaciones alimentarias que obedecen a la consigna de “quién come a quién” entre las distintas poblaciones.
 |
En otras palabras, las cadenas alimentariasindican qué seres vivos se alimentan de otros que habitan el mismo ecosistema.
Estas relaciones que se establecen entre los diversos organismos en su ambiente natural tienen dos consecuencias de gran importancia: el flujo de energía y la circulación de la materia.
Flujo de energía
Este flujo va desde los organismos autótrofos(por lo general, organismos que realizanfotosíntesis) hacia otros que se alimentan de ellos y que corresponden a herbívoros. A su vez, los herbívoros son presas de otros animales: losdepredadores. Se constituye así una verdadera cadena para la vida, donde cada eslabón corresponde a un ser vivo.
Circulación de materia
Ésta se traspasa de eslabón a eslabón en la cadena alimentaria, a través de las interacciones que se establecen entre los organismos que la conforman.
Aquí ya podemos definir en propiedad una cadena alimentaria, y podemos decir que es aquella sucesión en la cual las agrupaciones de organismos (cada uno representando un eslabón) establecen interacciones de manera tal que los primeros son alimento de los segundos traspasándose sucesivamente materia y energía de un eslabón al siguiente.
El primer eslabón, o primer nivel trófico, de cualquier cadena alimentaria siempre está representado por los productores, organismos autótrofos, los vegetales, que son capaces de transformar la energía lumínica del Sol en un tipo de energía que puede ser utilizado por plantas, bacterias, animales, etc.
Entonces, podemos decir que productores son aquellos organismos fotosintéticos que “producen” energía útil para todos los seres vivos. La vida en el planeta se mantiene en una cadena alimentaria, gracias a estos organismos fotosintéticos.
El segundo eslabón, o segundo nivel trófico, lo ocupan los consumidores, organismos incapaces de utilizar la energía lumínica del Sol, y que para conseguir la energía necesaria para vivir deben alimentarse de otros organismos.
A los consumidores se les denomina heterótrofos, ya que el término significa: hetero = otro, diferente y trofos = alimentación.
Se distinguen diferentes tipos de consumidores, según sea el nivel de la cadena en que aparecen.
Consumidores primarios o de primer orden son los organismos que se alimentan directamente de los productores.
Consumidores secundarios o de segundo orden son los organismos que se alimentan de los consumidores primarios.
En general, el nombre de los consumidores estará determinado por el nivel trófico en que aparezcan. Sin embargo, no es posible encontrar cadenas con más de cinco niveles, porque la cantidad de energía que se va traspasando de un nivel trófico al siguiente va disminuyendo de manera importante.
 |
| Red trófica terrestre. |
Otro grupo de organismos que son de gran relevancia para el flujo normal de materia y energía, a través de una cadena alimentaria, son los denominados descomponedores.
Descomponedores son los microorganismos que habitan en el suelo y son los encargados de degradar y descomponer organismos muertos o restos de ellos.
Ejemplo de descomponedores son los hongos y lasbacterias.
Esto determina que la materia que formaba parte de los seres vivos sea "devuelta" al ambiente, específicamente al suelo, donde puede volver a ser utilizada por otros organismos como los productores, los que a su vez los transmitirán a los consumidores de primer orden y así sucesivamente a lo largo de la cadena. El hecho de que los descomponedores actúen sobre restos de organismos muertos puede hacer pensar que siempre actúan en el último nivel trófico. Sin embargo, los descomponedores pueden actuar en cualquier nivel trófico.
En la naturaleza, sin embargo, no se da el hecho de que un consumidor primario se alimente sólo de un tipo específico de planta, o que un consumidor secundario se alimente sólo de un tipo de presa.
En realidad, las poblaciones establecen interacciones de alimentación o interacciones tróficas, bastante más complejas que lo que representa una cadena.
Se habla de Redes tróficas o Redes alimentarias para señalar un conjunto de cadenas que se interconectan en algunos niveles tróficos. De esta forma, un productor, como la hierba de un prado, puede ser pastoreado por más de un herbívoro o consumidor primario, como, por ejemplo, una cabra, una vaca, un conejo, etc.; a su vez, la cabra, lo mismo que la vaca, puede ser presa para dos o más consumidores secundarios. Se aprecia entonces lo difícil que es representar estas complejas interacciones en forma lineal. Más bien se obtiene una malla de flechas que sugieren el flujo de materia y energía, que se da entre las poblaciones interactuando entre sí.
Las redes tróficas corresponden a la representación de varias cadenas, que se interconectan en diferentes niveles alimenticios.
Fuente: http://www.profesorenlinea.cl/ecologiaambiente/Cadenas_alimentarias_o_troficas.html
HOMEWORK:
Consulta como esta compuesta una cadena trofica acuatica y dibujala
4° PERIODO

SEMANA DEL 24 AL 28 DE AGOSTO


::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::
SEMANA EL 17 AL 21 DE AGOSTO
SEMANA DEL 24 AL 28 DE AGOSTO
| Estructura de un ecosistema | |
| Flujo de energía
De toda la energía solar que llega a la superficie terrestre, sólo una pequeña parte, entre un 0,1% y 1% se incorpora a los organismos productores o autótrofos.
A partir de esta entrada de energía solar comienza un flujo unidireccional de energía a través de todos los organismos de un ecosistema, que fluye desde los organismos autótrofos hasta los heterótrofos, hasta que finalmente se disipa en el medio ambiente.
|
El Sol, fuente básica de energía
Casi toda la energía de que disponemos proviene del Sol. Él es la causa de los vientos, de la evaporación de las aguas superficiales, de la formación de nubes, de las lluvias y, por consiguiente, de los saltos de agua. Su calor y su luz son la base de numerosas reacciones químicas indispensables para el desarrollo de los vegetales y de los animales que con el paso de los siglos han originado combustibles fósiles como el carbón o el petróleo. La radiación solar tiene otra importancia capital: otras formas de energía renovable, como el viento, las olas o la biomasa no son más que manifestaciones indirectas de ésta.
::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::
SEMANA EL 17 AL 21 DE AGOSTO
HOLA CHICOS, EL DÍA 26 DE AGOSTO CORRESPONDE EL EXAMEN DE BIOLOGÍA.
LOS TEMAS QUE SE INCLUIRÁN EN EL EXAMEN SON:
La REPRODUCCIÓN
la REPRODUCCIÓN sexual: gametogenesis y FECUNDACIÓN.
Adaptaciones SEXUALES animales .
tipos de desarrollo embrionario.
la REPRODUCCIÓN humana: aparatos reproductores masculinos y femeninos, la madurez sexual en hombres y en mujeres
el ciclo menstrual
cuidados del sistema reproductivo
SEMANA DEL 17 AL 21 E AGOSTO
¿Qué es el ciclo menstrual?
Todos los meses, el cuerpo de la mujer se prepara para un eventual embarazo. A esta preparación se la llama “ciclo menstrual”, la sangre menstrual es sólo una parte de este ciclo.
El ciclo menstrual, puede durar entre 23 y 35 días, el promedio es de 28. Se cuenta como día 1 del ciclo al día en el que comienza la menstruación, aunque sólo sean unas gotas. El ciclo termina con el inicio de la siguiente menstruación.
Es importante tener en cuenta que estos plazos no son fijos ni estables, porque el cuerpo no es una computadora ni una máquina.

¿Qué sucede durante el ciclo menstrual?
Durante la primera mitad del ciclo menstrual, los niveles de la hormona llamada estrógeno aumentan y hacen que la pared interna del útero (que se llama endometrio), crezca y se ensanche. Al mismo tiempo, por el estímulo de diferentes hormonas, el óvulo empieza a madurar en uno de los ovarios. En la mitad del ciclo, otra hormona hace que el óvulo abandone el ovario. A esto se le llama ovulación.
La ovulación es la salida de un óvulo maduro de uno de los ovarios de la mujer. Se produce en la mitad del ciclo, entre menstruación y menstruación. En un ciclo de 28 días la ovulación se da en el día 14 aproximadamente. El óvulo es expulsado por el ovario y absorbido por la trompa de Falopio, por donde viaja seis días y medio hasta llegar al útero.
Durante la segunda mitad del ciclo menstrual, mientras el útero se sigue preparando para un posible embarazo, el óvulo comienza a desplazarse a través de la trompa de Falopio hacia el útero. Si un espermatozoide se encuentra con el óvulo en las trompas, lo fertiliza, y luego el óvulo se adhiere a la pared del útero, la mujer queda embarazada.
Si el óvulo no es fertilizado o no se adhiere, se disuelve o es absorbido por el cuerpo. Como el embarazo no se produjo, los niveles de hormonas bajan y el recubrimiento ensanchado del útero (el endometrio con el que empezó esta explicación) es liberado en forma de sangre menstrual.
¿Qué es la menstruación?
La menstruación es el sangrado mensual de la mujer. También se le llama regla, período menstrual, días húmedos, “esos días”, “Andrés” y, algunas personas le dicen “estar enferma” o “enfermedad” (aunque sea un proceso natural del cuerpo). En realidad, la sangre menstrual es en parte sangre y en parte tejido del endometrio que se desprende porque no albergó a ningún óvulo fecundado. Fluye desde el útero y sale del cuerpo a través de la vagina. La mayor parte de los períodos menstruales duran entre tres y siete días. Pueden no ser igual todos los meses, ni ser iguales a los de otras mujeres.
-
SEMANA DEL 10 AL 14 DE AGOSTOVÍDEO RELACIONADO:
Órganos sexuales femeninos
Monte Venus
Es una prominencia de tejido graso que se forma bajo la piel de la región del pubis femenino. La piel en esta zona durante la pubertad se vuelve más oscura y se cubre de vello.
Clítoris
Es un órgano pequeño y eréctil, es decir que aumenta de tamaño por la llegada de sangre a esa zona durante la excitación sexual, mide desde 1 a 3 cm., tiene forma cilíndrica y se encuentra cubierto por el capuchón del clítoris que es la continuación de los labios menores. Este órgano presenta infinidad de terminaciones nerviosas que proveen de sensaciones sexualmente placenteras.
Labios Mayores
Son dos pliegues de piel de aproximadamente 7 cm. de largo. La piel que los cubre es más oscura que la del resto del cuerpo, en el exterior están cubiertos de vello y poseen glándulas que empiezan a secretar sudor (sudoríparas) y grasa (sebáceas) en la pubertad. Poseen una gran sensibilidad.
Labios Menores
Son dos pliegues de piel que rodean el orificio vaginal, el meato urinario y terminan formando el capuchón del clítoris. Estos pliegues que crecen en la pubertad, cuentan con un tejido eréctil que les permite aumentar de volumen durante la excitación sexual y poseen gran sensibilidad.
Himen
Es una estructura que se encuentra en el interior de la entrada de la vagina. Es una membrana que cierra parcialmente la entrada de la vagina en algunas de las mujeres que no han tenido relaciones sexuales con penetración. Existen mujeres que nacen sin himen, en otras es elástico, conocido como “himen complaciente”, que no se rompe durante las relaciones sexuales con penetración. La mayoría suele romperse durante la primera relación sexual con penetración, produciéndose un pequeño sangrado. La ausencia de sangrando durante el primer contacto sexual, o la ausencia de himen en algunas mujeres de ninguna manera representan una prueba de que la mujer no sea virgen.
Vagina
Este órgano está hecho de un tejido muy elástico y sus paredes están adheridas, como si fuera un globo desinflado. Durante la excitación sexual, cierta cantidad de líquido que proviene de la sangre atraviesa las paredes vaginales produciéndose así la lubricación vaginal. La vagina también es la vía por donde sale el sangrado que viene del útero durante la menstruación. La vagina es tan flexible que durante el parto el bebé pasa por la vagina, también se le conoce como “canal de nacimiento”. A la entrada de la vagina se le denomina “orificio vaginal”.
Glándulas de Bartholin
Son dos glándulas ubicadas a cada lado del orificio vaginal, cada una de ellas secreta durante la excitación sexual gotas de una sustancia acuosa, que contribuyen a la lubricación vaginal.

Trompas de falopio o tubas uterinas
Son dos tubos de comunicación entre el ovario y el útero, su función es la de transportar al óvulo desde el ovario en dirección al útero. Las trompas en su interior están recubiertas por pequeñas vellosidades, las fimbrias, que crean corrientes hacia el útero y así se lleva a cabo la transportación. En el interior de las trompas se lleva a cabo la fecundación, es decir, la unión de un espermatozoide con un óvulo.
Útero o Matriz
Es un órgano hueco en forma de pera invertida inclinada ligeramente hacia el frente. Su función es dar albergue al huevo fecundado durante todo su desarrollo, es decir que en el útero se lleva a cabo el embarazo. El útero está constituido de tres capas:
Endometrio
Es la capa mucosa interna, con gran cantidad de vasos sanguíneos, susceptible a los cambios cíclicos hormonales que se desprende durante la menstruación para volver a crecer en el siguiente ciclo.
Cuello del útero o cerviz
Es la capa mucosa interna, con gran cantidad de vasos sanguíneos, susceptible a los cambios cíclicos hormonales que se desprende durante la menstruación para volver a crecer en el siguiente ciclo.
Ovarios
Son dos cuerpos en forma de almendra que se encuentran sostenidos por un ligamento y a cada lado del útero o matriz. Tienen una doble función, por un lado son los responsables de la maduración de los óvulos y, por el otro, de la producción de hormonas.

REPRODUCCIÓN HUMANA






SEMANA DEL 27 AL 31 DE JULIO
HOLA CHICOS
ESTA SEMANA VEREMOS COMO SE DESARROLLAN LOS EMBRIONES DE DIFERENTES ESPECIES.

::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::::
SEMANA DEL 20 AL 24 DE JULIO



SEMANA DEL 13 AL 17 DE JULIO




semana del 6 al 10 de julio
RECORDEMOS QUE PARA QUE PUEDAN HEREDARSE LOS CARACTERES EN ORGANISMOS EUCARIOTAS DEBE EXISTIR LA REPRODUCCIÓN. EN EL CASO PARTICULAR DE LOS MAMÍFEROS, LAS CÉLULAS SEXUALES DESEMPEÑAN UN PAPEL FUNDAMENTAL EN ESTE PROCESO. CUANDO LA REPRODUCCIÓN SE DA GRACIAS A LA UNIÓN DE LOS GAMETOS SEXUALES DECIMOS QUE ES UNA REPRODUCCIÓN SEXUAL.
LA REPRODUCCION SEXUAL Y SU PAPEL EN LA HERENCIA DE CARACTERES.

3° PERIODO
SEMANA DEL 9 AL 12 DE JUNIO
ESTA SEMANA HABLAREMOS DE COMO SE HEREDAN LOS CARACTERES GENÉTICOS. CON ESTE TEMA EMPEZAREMOS EL TERCER PERIODO POR LO TANTO DEJO INFORMACIÓN QUE SERA DE AYUDA PARA EMPEZAR.
Las características de los seres vivos se heredan a partir de la molécula de ADN quien es la única biomolécula capáz de almacenar y transmitir los caracteres heredables de tipo Fenotipo o visibles y Genotipo o recesivos a la descendencia.
Cuando el ADN se duplica semiconservativamente las moléculas de ADN hijas quienes contienen la información genética de los antecesores transmiten la misma información hereditaria a la nueva descendencia.
En organismo unicelulares procariotas ( bacterias) el intercambio de información genética se realiza por Conjugación, en organismo multicelulares eucariotas la información genética esta contenida en los GENES almacenados en cuerpos compáctos llamados Cromosomas en unas cavidades especializadas para alojar a los genes llamadas Locus o Losci, cada gen es portador de una determinada información genética visible o recesiva.
Por fecundación cuando el óvulo e une con el espermatozoide se origina la célula huevo o cigoto y se reconstituye el núcleo diploide de la misma, este núcleo diploide contiene la mitad de la información genética aportada por el pronúcleo femenino y la otra mitad aportada por el pronúcleo masculino, luego por los mecanismos de la herencia( 3 leyes de Mendel) se establecen cruzas entre individuos de razas puras ( homocigotas) y heterocigotas para la perpetuación de las especies en el tiempo y transmisión de caracteres biológicos hereditarios.
SEMANA DEL 1 AL 5 DE JUNIO
HOLA CHICOS, ESTA SEMANA CORRESPONDE EL EXAMEN DE BIOLOGÍA, MAS EXACTAMENTE EL DÍA JUEVES 4 DE JUNIO, LOS TEMAS ASIGNADOS SON:
GENÉTICA MENDELIANA.
QUIEN FUE GREGOR MENDEL Y EN QUE CONSISTIERON SUS TRABAJOS.
LAS TRES LEYES DE MENDEL.
EJERCICIOS DE GENÉTICA USANDO CUADROS DE PUNNET.
CODOMINANCIA Y EJERCICIOS DE HERENCIA DE LOS GRUPOS SANGUÍNEOS
HERENCIA LIGADA AL SEXO.
______________________________________________________________________________________________________
semana del 25 al 29 de mayo
Esta semana veremos el tema de herencia ligada al sexo
La especie humana posee 46 cromosomas dispuestos en 23 pares, de esos 23 pares 22 son somáticos o autosomas (heredan caracteres no sexuales) y uno es una pareja de cromosomas sexuales (llamados también heterocromosomas ogonosomas), identificados como XX en las mujeres y como XY en los hombres.
Esta pareja de cromosomas sexuales no solo llevan los genes que determinan el sexo, sino que también llevan otros que influyen sobre ciertos caracteres hereditarios no relacionados con el sexo.
Ver: Herencia y genética
Hay caracteres que sin ser caracteres sexuales primarios (órganos genitales, gónadas) o secundarios (barba del hombre, pechos de las mujeres), solo aparecen en uno de los dos sexos, o si aparecen en los dos, en uno de ellos son mucho más frecuentes.
A estos caracteres se les denomina caracteres ligados al sexo.
Variantes en la herencia ligada al sexo
Vimos que los cromosomas sexuales constituyen un par de homólogos (XX en la mujer y XY en el hombre); sin embargo, en el par XY un segmento de cada cromosoma presenta genes particulares y exclusivos (segmento heterólogo, llamado también diferencial o no homólogo), la porción restante de los cromosomas del par XY corresponde al sector homólogo, como se grafica en el esquema siguiente:
 |
Los varones sólo llevan un representante de cada gen ubicado en el sector heterólogo del X (en tanto poseen un X) y las mujeres portan dichos genes por pares (en tanto poseen dos X). Por consiguiente, la transmisión y expresión de estos genes dependen del sexo de los individuos.
Entonces, la herencia ligada al sexo se refiere a la transmisión y expresión, en los diferentes sexos, de los genes que se encuentran en el sector no homólogo (heterólogo) del cromosoma X heredado del padre.
También podemos decir que la herencia ligada al sexo no es más que la expresión en la descendencia de los genes ubicados en aquellas regiones del cromosoma X que no tienen su correspondencia en el cromosoma Y.
En el sexo femenino, la presencia de dos cromosomas X hace que los genes contenidos en estos se comporten como si se encontraran en autosomas, con normalidad.
 |
| Daltonismo y hemofilia, dos anomalías recesivas ubicadas en el segmento diferencial del cromosoma X. |
Así, pues, un carácter determinado por un gen del cromosoma X aparecerá si la mujer tiene un alelo dominante en cada uno de estos cromosomas, o si tiene dos alelos recesivos, uno en cada uno de ellos (homcigota en ambos casos).
Si, en cambio, la mujer es heterocigota para ese carácter, igual se manifestará el alelo dominante. Es decir, se trata de un modelo de herencia clásico y normal, comparable a los mencionados en las leyes de Mendel.
El caso del hombre es radicalmente distinto. Si los genes se encuentran en la zona del cromosoma X que tiene su parte correspondiente (homóloga) en el Y, actúan como en el caso anterior.
De hecho, se han encontrado algunos alelos en los sectores homólogos de los cromosomas XY (llamados genes pseudoautosómicos) y dadas las características de estas regiones especiales de los cromosomas sexuales, los trastornos producidos a causa de estos genes se heredarán de acuerdo a las leyes genéticas que rigen la herencia autosómica (o sea, funcionan igual que los autosómicos).
Pero es mucho más frecuente que los genes estén en una parte del cromosoma X que no tenga correspondencia en el Y. Si esto sucede, los alelos se manifestarán siempre, ya sean dominantes o recesivos.
Por tanto, debemos recordar que los genes ligados a los cromosomas sexuales pueden ser tanto recesivos como dominantes.
Herencia recesiva ligada al sexo
En los hombres, los cromosomas X e Y se pueden aparear durante la meiosis (en la Profase I) por sus fragmentos homólogos y por tanto se pueden producir entrecruzamientos en esta zona.
Herencia ligada al cromosoma Y o herencia holándrica
Todos los genes que se encuentran en el segmento diferencial o no homólogo del cromosoma Y son heredados únicamente por los hijos varones y se manifestarán en todos los hombres que los lleven y sólo en los hombres, independientemente de que sean dominantes o recesivos.
La transmisión de los genes situados en el segmento no homólogo del cromosoma Y es la herencia holándrica.
Herencia ligada al cromosoma X.
La herencia ligada al cromosoma X quiere decir que el gen que causa el rasgo o el trastorno se localiza en el cromosoma X .
Cabe recordar que las mujeres poseen dos cromosomas X mientras que los hombres poseen un cromosoma X y un cromosoma Y. Los genes del cromosoma X pueden serrecesivos o dominantes, y su expresión en las mujeres y en los hombres no es la misma debido a que los genes del cromosoma Y no van apareados exactamente con los genes del X.
Los genes recesivos ligados al cromosoma X se expresan en las mujeres únicamente si existen dos copias del gen (una en cada cromosoma X). Sin embargo, en los varones sólo debe haber una copia de un gen recesivo ligado al cromosoma X para que el rasgo o el trastorno se exprese.
Por ejemplo, una mujer puede ser portadora de un gen recesivo en uno de sus cromosomas X sin saberlo y transmitírselo a su hijo, que expresará el rasgo o el trastorno.
Entre los ejemplos de trastosnos recesivos ligados al cromosoma X se destacan los casos del daltonismo y la hemofilia, enfermedades provocadas por un gen recesivosituado precisamente en el segmento diferencial del cromosoma X.
Viernes 21 del 2015
Hola
chicos la actividad que deben presentar el día lunes 25 de mayo, es escoger del siguiente cuadro 3 características y realizar las observaciones en padres, hermanos y usted mismo, para determinar los posibles genotipos de cada uno. recuerden que deben presentar el diagrama de distribución de los alelos como les explique en clases. RECUERDA REPRESENTAR CON LETRAS MAYÚSCULAS LOS ALELOS DOMINANTES Y CON MINÚSCULAS LOS RECESIVOS.
Bendiciones


SEMANA DEL 19 AL 22 DE MAYO





VÍDEO HERENCIA GENÉTICA HUMANA
HOMEWORK
A PARTIR DEL SIGUIENTE CUADRO ESCOGE TRES CARACTERÍSTICAS, LAS CUALES OBSERVARAS EN TU FAMILIA Y A PARTIR DE LA INFORMACIÓN DE CUALES SON DOMINANTES Y REPRESIVOS INTERPRETA TUS CARACTERÍSTICAS, LAS DE TUS PADRES Y HERMANOS, Y SI PUEDES LA DE TUS ABUELOS PATERNOS Y MATERNOS, Y A PARTIR DE AHÍ ELABORA UN ÁRBOL GENEALÓGICO O LINEAS CON LOS POSIBLES GENOTIPOS SEGÚN SEA EL CARÁCTER EVALUADO.

Semana del 11 al 14 de mayo

chicos
Esta semana corresponde ver el tema de herencia de los grupos sanguíneos. por lo cual dejo material para que se preparen con tiempo para la clase.



vídeos relacionados con la temática
HOMEWORK
CONSULTAR QUE ES EL FACTOR RH Y ESTUDIARLO PARA SUSTENTARLO EN LA PRÓXIMA CLASE
______________________________________________________________________________________________________
HOMEWORK
REALIZA UN TRABAJO ACERCA DE QUIEN FUE GREGOR MENDEL, EN QUE CONSISTIERON SUS TRABAJOS, QUE LOGRO CON ELLOS Y QUE RELEVANCIA TIENE EN LA GENÉTICA ACTUAL.
ENTREGA PARA EL 20 DE ABRIL DEL 2015
____________________________________________________________
____________________________________________________________
HOLA CHICOS ESTA ES LA TEMÁTICA CON LA QUE INICIAMOS EN EL SEGUNDO PERIODO.
MUCHOS ÉXITOS EN ESTA NUEVA ETAPA DEL AÑO ESCOLAR.
2 DE MARZO DEL 2015
TEMA: DIVERSIDAD GENÉTICA
HOLA CHICOS POR AQUÍ LES DEJO LA PRESENTACIÓN PPT DE LA CLASE DE DIVERSIDAD GENÉTICA CON LA QUE SE TRABAJO EN EL AULA.
_________________________________________________________________________________
HOLA. CONTINUANDO CON EL TEMA DE: COMO INFLUYE EL MEDIO AMBIENTE EN LOS SERES VIVOS LES DEJO LA PRESENTACIÓN PPT VISTA EN CLASES, ACERCA DE LAS ADAPTACIONES DE LOS SERES VIVOS
Miss Maria Tereza Carmona
Biology teacher.
RECOMBINACIÓN GENÉTICA
HOLA CHICOS, LA PRÓXIMA SEMANA VEREMOS EL TEMA DE RECOMBINACIÓN GENÉTICA. ESTE SERA EL VÍDEO A PROYECTAR EN LA CLASE. PUEDEN IR REALIZANDO INVESTIGACIÓN INDEPENDIENTE ACERCA DE LA TEMÁTICA.
ATT: Miss Mayte Carmona
_________________________________________________________________________________
RELACIONES RECIPROCAS ENTRE ESPECIES

________________________________________________________________________________
_________________________________________________________________________
HOLA QUERIDOS CHICOS DEL GRADO 10°.
Como les comente en el desarrollo de la clase de biología, les facilito el material para reforzar el tema visto en clases (ADN, estructura y funciones). deben observarlo para socializarlo en la próxima clase, ademas les recomiendo anotar cualquier inquietud respecto al tema, de tal manera que me puedan exponer sus dudas y así poder trabajarlas juntos, dando así solución a estas.
Miss Maria Tereza Carmona
Biology teacher.
